The secret world in the gaps between brain cells
DOI: 10.1063/PT.3.4999

A map of the US with only the state boundaries shown appears fairly blank, but zooming in to a region of a large city shows a complicated collection of buildings delineated by streets. The brain turns out to be similar—deceptively simple until you look closely.
Given its extraordinary properties, brain tissue is surprisingly unprepossessing. The human cerebral cortex appears to be made of a whitish substance about as featureless as blancmange. Its monotony is broken only by the folding of the cortical surface that enables more brain tissue to pack in the skull. The same featurelessness holds true for a small sample of brain tissue when removed and viewed under a light microscope. But if appropriate dyes are applied, vast numbers of distinct cells appear.
Beginning in the 19th century and continuing through today, light microscopists have focused on the shapes of and connections between brain cells and given little thought to the spaces between them. The advent of electron microscopy finally supplied the necessary resolution to discern how cells were stacked together, as shown in figure
Figure 1.

Brain tissue is more complicated than it appears at the macroscopic level. The brain’s apparent featurelessness hides a complicated microscopic structure of brain cells (grayscale) and extracellular space (false colored in blue) between those cells, shown here from a transmission electron micrograph of an adult mouse’s cerebral cortex that has been cryo-processed to preserve the spaces between cells. (Courtesy of Graham Knott, Swiss Federal Institute of Technology, Lausanne; adapted from ref.

It’s now known that every cell in the brain is separated from its neighbor by a fluid-filled extracellular space (ECS), which forms sheets and tunnels, as shown on

Courtesy of Terrence Sejnowski, Salk Institute for Biological Studies; adapted from T. J. Sejnowski, “Nanoconnectomics,” in Micro-, Meso- and Macro-Connectomics of the Brain, H. Kennedy, D. Van Essen, Y. Christen, eds., Springer (2016), p. 4, under license CC BY-NC 2.5 .

Invisible ECS
Evidence for a substantial ECS emerged slowly. Early electron microscopy measurements suggested little or no gaps between brain cells. Then in the 1960s some heretical microscopists, such as Anthonie Van Harreveld of Caltech, argued that the procedures used by most microscopists to prepare brain tissue for observation caused the ECS to collapse. They showed that if the tissue was instead rapidly frozen, gaps could be seen between the cells. 1
Another limitation of electron microscopy images is that they’re taken on dead tissue. Recognizing that drawback, other researchers in the 1960s used live tissue injected with solutions of select radiotracers that were not expected to enter cells. A radiotracer, or radioactive tracer, is a chemical compound with one or more atoms replaced by a radioisotope so that the tracer’s position or concentration can be monitored by its radioactive decay. Those tracers penetrated brain tissue, which hinted that there must be spaces between cells. But the initial results were ambiguous as to what space the tracers were moving in.
Getting quantitative results from radiotracers required work from such pioneers as Joseph Fenstermacher of the National Cancer Institute in Bethesda, Maryland, and Clifford Patlak of the National Institute of Mental Health, also in Bethesda.
2–4
In the 1970s, they combined physiological expertise with quantitative analysis to understand the diffusion of cell-impermeant tracers, such as radiolabeled sucrose, into brain tissue. By fitting a solution to the diffusion equation to experimental spatial concentration profiles, they extracted the radiotracer’s effective diffusion coefficient
The effective diffusion coefficient was always smaller than the free diffusion coefficient in water
Many of the limitations of the radiotracer method were overcome with the introduction of two techniques: the real-time iontophoresis (RTI) method in 1980 and the integrated optical imaging (IOI) method in 1993. My research group developed those techniques, which are often referred to as examples of a point-source paradigm. 2 , 4 , 5 Soon after the introduction of the RTI method, Eva Syková at the Czech Academy of Sciences in Prague adopted the technique and has since applied it to clinically important problems, including ischemic stroke, brain tumors, and aging. 2
To the point
In the RTI method, a micropipette and an ion-selective microelectrode with micrometers-wide tips are inserted in living brain tissue.
2
,
4
,
5
A current passes through the micropipette, which contains an ionic solution of a chosen small probe molecule that stays in the ECS, and a precise number of cations or anions are released into the brain, a process known as iontophoresis. The probe molecules then diffuse through the ECS, and the concentration as a function of time is measured about 100 µm away by the ion-selective microelectrode (see the
Quantifying the ECS
Two diffusion-based measurement techniques, real-time iontophoresis (RTI) and integrated optical imaging (IOI), explore the extracellular space in an anesthetized rat or mouse. In the RTI method, a source micropipette releases a burst of small ions, typically the cation tetramethylammonium (TMA), at a depth of about 200 μm below the surface of the brain. An ion-selective microelectrode positioned about 100 µm away then detects the diffusing molecules. The micropipette and microelectrode can alternatively be inserted parallel to one another to access almost any region of the brain.
The concentration

For the IOI method, only the source micropipette is required. A brief applied pressure pulse releases fluorescent molecules in the ECS, which gradually spread. An overhead microscope and CCD (top of image) track that spread over time. The fluorescence intensity as a function of distance is fitted with a solution to the diffusion equation—in this case, a Gaussian curve—and
The RTI method gives results in minutes, which enables the recording of dynamic changes in ECS properties through repeated measurements. Another major advantage of the RTI method is that it can reveal simultaneously
The hindrance to diffusion is characterized by a dimensionless parameter known as the tortuosity
The RTI method is constrained by the availability of microelectrodes that are sensitive to molecular probes of interest. So far, they can be fabricated to sense only a few specific small molecules, and overwhelmingly, the method has used the cation tetramethylammonium, with a molecular mass of 74. Because they’re so small, those molecules approximate a point in space and are thus valuable for mapping geometry, but many molecules of biological interest are much more spatially extended.
About a decade after the first implementation of the RTI method, researchers extended the point-source paradigm to macromolecules with the introduction of the IOI method.
2
,
3
Again, a micropipette released molecules, but those molecules had fluorescent markers on them so that the distribution could be imaged by optical microscopy. The intensity distribution, representing the concentration of fluorescent molecules, could then be measured and fitted to a solution of the diffusion equation to give the tortuosity averaged over about 100 µm (see the
Although the point-source paradigm was successful, the spatial resolution of the RTI and IOI methods was, at best, tens of micrometers, and electron microscopy and macromolecular-diffusion behavior indicated that the ECS was tens of nanometers wide. Now, an era of discovery is on the horizon with the introduction of techniques that rely on superresolution optical microscopy with a resolution of 50–150 nm.
One such technique is superresolution shadow imaging (SUSHI), developed in the laboratory of Valentin Nägerl of the University of Bordeaux in France. 7 The SUSHI technique is based on stimulated-emission-depletion microscopy. In that technique, some of the fluorophores deposited on a sample are temporarily deactivated so that when a laser spot shines on them, only a small central area lights up and thus the resolution is improved beyond the diffraction limit. In the SUSHI method, the fluorophores are administered as a dye solution that permeates the ECS, and the method images the local volume occupied by them.
Laurent Cognet’s group at the University of Bordeaux proposed another approach that tracks carbon nanotubes more than 100 nm long and about 1 nm wide.
8
Those nanotubes have an optical response in the near-IR and offer a spatial resolution of about 50 nm. The probes are tracked as they wander in the ECS, and those results measure the diffusion properties and the ECS’s local structure. The nanotube technique and the SUSHI method have revealed that the ECS structure is very heterogeneous, a conclusion supported by recent advances in electron microscopy on rapidly cooled tissue, such as in figure
Physics of diffusion
The key physical concept underlying the radiotracer method, the point-source paradigm, and single-particle tracking is diffusion. The diffusion of molecular signals, metabolites, and drugs in the brain is vitally important for the brain’s normal functioning and for medical interventions. It’s also vital for understanding how specific probe molecules reveal structural information about the ECS. And underlying diffusion is the random walk.
Water’s concentration in a beaker or in the interstitial fluid of the ECS is about 50 moles per liter, whereas small probe molecules are introduced at a concentration of millimoles per liter or less. At the temperature of the human body, about 37 °C, such probe molecules move because of thermal energy and collide constantly with water molecules and occasionally with the cell walls bounding the ECS. Each collision sends the probe in a random direction until it’s redirected by the next collision; that ever-changing trajectory is a random walk. Given enough walkers and sufficient time, random walks explore the whole of a domain. And if the fruits of that exploration can be analyzed, then the structure in which the molecules move becomes apparent, as shown in figures
Figure 2.

Random walks explain how probes diffuse through brains. In these simulated two-dimensional random walks, 50 particles start at the center of each image and take 1500 steps of equal size but in random directions either (a) unimpeded or (b) surrounded by circular obstructions that represent cells. Each of the 50 particle trajectories was assigned one of six colors. The particles spread farther in the absence of hindrance.

With the help of superresolution optics aided by computation, emerging single-particle tracking methods follow one probe particle moving through the ECS to yield unprecedented local detail, as shown in figure
Figure 3.

Single-particle tracking and superresolution imaging reveal the local structure of the extracellular space. (a) Single-walled carbon nanotubes diffuse through a living slice of an adult mouse’s brain with the instantaneous diffusion coefficients relative to unimpeded diffusion (
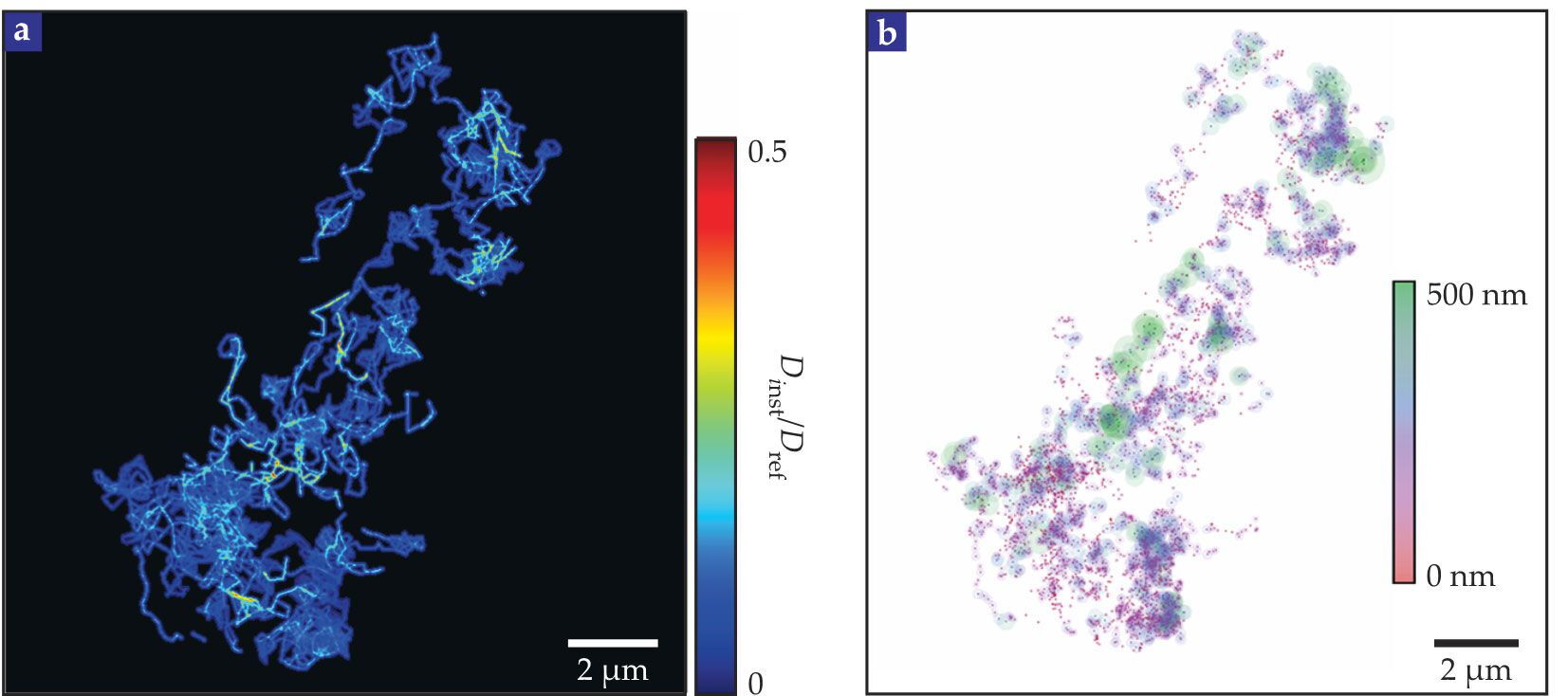
By contrast, in the point-source paradigm, typically about 1014 probe molecules are injected into the brain, and each one sets off on a random walk. The concentration distribution measures the aggregate of all those evolving trajectories and can be analyzed on a macroscopic scale by solving the diffusion equation, which is a partial differential equation otherwise known as Fick’s second law (see the box
After a population of molecules is released from a point and undergoes subsequent random walks, the molecules’ distances
Does advection contribute?
Since the radiotracer experiments of the 1960s, researchers have accepted that diffusion takes place in the ECS. But from time to time, the idea surfaced that advection—that is, the transport of substances by bulk flow of the interstitial fluid filling the ECS—might also occur. Important evidence for advection came around 1980 from Helen Cserr of Brown University. She performed a different type of tracer experiment that extended over longer time periods and over multiple brain compartments. She showed that injected radiotracers of different molecular weights and sizes cleared from the brain at the same rate.
The physics of diffusion and advection are different. Diffusion from a point source is rapid in the vicinity of the source but quickly diminishes with distance, and the travel time depends on the size of the molecule. Flow-mediated advection, on the other hand, maintains speed and can carry material over long distances. Cserr argued that the weight-independent clearing rate in the ECS indicated that the main transport mechanism was some overall flow of the interstitial fluid that carried all radiotracers with it. If the mechanism was diffusion, on the other hand, then the tracers would have cleared at distinct rates related to their molecular sizes. 2 But she and others couldn’t directly show such bulk flow or advection.
The possibility of flow in the ECS has since received further support. Recent work suggested that the flow originates in the perivascular spaces, which form a sheath around the many blood vessels that penetrate the brain. Cerebrospinal fluid enters the sheaths surrounding arteries near the brain surface and then moves into the brain. The proposed idea is that some of that fluid leaves the perivascular spaces and moves through the ECS to exit at the perivascular space around veins. Flow in the perivascular space has been established, but flow in the ECS is not yet confirmed. 10
Maiken Nedergaard of the University of Rochester in New York proposed that ECS flow pattern, which she called the glymphatic system, 10 in 2012. She speculated that the glymphatic system may flush waste material out of brain tissue. Such waste material includes amyloid-beta protein, which is thought to play a role in the development of Alzheimer’s disease.
Lessons learned
In 1979 I drew an illustration, shown in figure
Figure 4.

The extracellular space (ECS) looks different at various length scales. (a) Depicted logarithmically from high magnification (left) to low magnification (right), the ECS evolves from tendrils of extracellular-matrix molecules to the cell membrane’s lipid bilayer to whole cells. (Adapted from ref.

On the left, figure
At the magnification of a few micrometers, electron microscopy, shown in figure
At the 10–100 µm scale, diffusion measurements from the analysis of probe molecules become relevant. Radiotracers provided a preliminary estimate that the ECS occupied 15–20% of brain tissue volume. The RTI method confirmed that result and found similar values for diverse brain regions in rats, mice, humans, some nonmammalian vertebrates, and even octopuses. 2 The ECS volume fraction is at least 20% in the sleeping or anesthetized brain but intriguingly seems to contract in the awake state 13 to around 15%. The surprisingly high volume of ECS can be understood by imagining that the outer membrane of every cell, including its extensions, is accompanied by a thin atmosphere of ECS and that any given volume of brain tissue has an enormous number of cells.
Why does brain tissue have so much ECS? One answer is that, in order to exchange electrical signals, nerve cells maintain a potential difference between their insides and outsides. That potential arises from a difference in ionic concentrations across the ion-selective cell membrane that creates a battery. So there must be a reservoir of ions external to the membrane, which is maintained by active transport of ions across cell membranes.
Another reason for the ECS is that substances need to diffuse between cells. Some of those substances, such as glucose, are involved in cellular metabolism, and some are waste products of metabolism. Those substances move to and from the vast network of blood vessels that permeate the brain. Other substances are signaling molecules that pass between cells. That chemical communication channel has long been discussed 14 and today is widely accepted and commonly called volume transmission. 11 , 15
Diffusion measurements also reveal that molecules are hindered as they move through the ECS,
2
as quantified by the tortuosity factor
The experimentally measured tortuosity suggested that dead spaces in the ECS cause additional delays to diffusing molecules.
16
Electron micrographs and superresolution optical images, such as those in figures
The IOI method showed that larger molecules are more hindered than smaller ones, probably because of increased interaction with the cell walls that bound the ECS. That assumption enabled an estimate of the ECS’s width, in addition to its relative volume. The estimate 3 was about 38–64 nm; by comparison, the cell body of a typical neuron is about 10–50 µm wide. As noted previously, the ECS is not uniform in size but has many enlargements or voids, which optical methods suggest can be several hundred nanometers wide. 7 , 8 So the estimated width is an average value.
Finally, dramatic changes in ECS volume have been shown to occur in pathological conditions, such as ischemia or stroke, when the local blood flow is cut off to part or all of the brain. The resulting lack of oxygen and glucose causes cells to rapidly swell and thus shrink the ECS volume fraction as some of the interstitial fluid’s salt enters cells and water follows suit to maintain osmotic balance. In that process, the ECS volume fraction drops as low as 5%, and the tortuosity increases 2 to around 2.0. Similar changes occur in what’s known as spreading depression or spreading depolarization—a condition thought to underly some types of migraine headache—when diffusion appears to briefly stop altogether. 6
A place for discovery
Just as the streets—with their constant streams of pedestrians, cars, and buses—are an essential element of a city, so too is the ECS an essential element of the brain. Streets define a city because they clearly delineate the buildings and allow the efficient exchange of people and goods between them. Similarly, the ECS permits the cells of the brain to maintain their identities and participate in intercellular molecular commerce. The ECS is a highly connected porous medium, so just like people in a well-designed city, molecules in the brain have many routes to move from one location to another.
Where cities and the ECS part company is that the layout of cities is largely two-dimensional, whereas the ECS is three-dimensional. Electron microscopy and superresolution optical methods provide only a 2D picture. A 3D electron-microscopy reconstruction of brain tissue exists, but it does not preserve the width of the ECS. To account for that limitation, a computer algorithm reinflated the reconstruction, as shown in figure
In addition to being 3D, the ECS is a dynamic structure that varies on multiple timescales. The change in volume fraction between sleeping and waking states shows variance sustained over hours, 13 while an analysis of spreading depression saw dramatic changes in volume fraction over tens of seconds or minutes. 6 Recently, rapid extracellular volume pulsations lasting a second or so have been detected during epileptiform activity. 18
History suggests that the present technical limitations are only temporary. The diffraction-limited resolution of light microcopy previously seemed insurmountable but is now beaten regularly. So too will present techniques likely be refined or replaced by the clever application of physical concepts, and with them will come new discoveries about the ECS’s interstitial content, traffic patterns, and its functional significance.
References
1. N. Korogod, C. C. H. Petersen, G. W. Knott, eLife 4, e05793 (2015). https://doi.org/10.7554/eLife.05793
2. E. Syková, C. Nicholson, Physiol. Rev. 88, 1277 (2008). https://doi.org/10.1152/physrev.00027.2007
3. D. J. Wolak, R. G. Thorne, Mol. Pharmaceutics 10, 1492 (2013). https://doi.org/10.1021/mp300495e
4. Y. Sun, X. Sun, Rev. Neurosci. 32, 363 (2021). https://doi.org/10.1515/revneuro-2020-0057
5. F. N. Soria et al., Front. Neurosci. 14, 570750 (2020). https://doi.org/10.3389/fnins.2020.570750
6. J. Hrabe, S. Hrabetova, Biophys. J. 117, 1783 (2019). https://doi.org/10.1016/j.bpj.2019.08.031
7. J. Tønnesen, V. V. G. K. Inavalli, U. V. Nägerl, Cell 172, 1108 (2018). https://doi.org/10.1016/j.cell.2018.02.007
8. C. Paviolo et al., Methods 174, 91 (2020). https://doi.org/10.1016/j.ymeth.2019.03.005
9. C. Nicholson, P. Kamali-Zare, Neurochem. Res. 45, 42 (2020). https://doi.org/10.1007/s11064-019-02793-6
10. H. Mestre, Y. Mori, M. Nedergaard, Trends Neurosci. 43, 458 (2020). https://doi.org/10.1016/j.tins.2020.04.003
11. L. Vargová, E. Syková, Philos. Trans. R. Soc. Lond. B Biol. Sci. 369, 20130608 (2014). https://doi.org/10.1098/rstb.2013.0608
12. J. W. Fawcett, T. Oohashi, T. Pizzorusso, Nat. Rev. Neurosci. 20, 451 (2019). https://doi.org/10.1038/s41583-019-0196-3
13. L. Xie et al., Science 342, 373 (2013). https://doi.org/10.1126/science.1241224
14. C. Nicholson, in The Neurosciences: Fourth Study Program, F. O. Schmitt, F. G. Worden, eds., MIT Press (1979), p. 457.
15. L. F. Agnati et al., eds., Volume Transmission Revisited, Elsevier (2000).
16. S. Hrabetova et al., J. Neurosci. 38, 9355 (2018). https://doi.org/10.1523/JNEUROSCI.1664-18.2018
17. J. P. Kinney et al., J. Comp. Neurol. 521, 448 (2013). https://doi.org/10.1002/cne.23181
18. R. Colbourn et al., J. Physiol. (London) 599, 3195 (2021). https://doi.org/10.1113/JP281544
More about the authors
Charles Nicholson is an emeritus professor in the department of neuroscience and physiology at the New York University Grossman School of Medicine in New York City.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}