Two Investigators of Pores in Cell Membranes Win Nobel Chemistry Prize
DOI: 10.1063/1.1650216
“For discoveries concerning channels in cell membranes,” as the Royal Swedish Academy of Sciences put it, two Americans have won this year’s Nobel Prize in Chemistry. Peter Agre of the Johns Hopkins School of Medicine in Baltimore earned his half of the prize for discovering water channels. Roderick MacKinnon of the Rockefeller University in New York City earned his half for structural and mechanistic studies of ion channels.
Water channels span cell membranes in our kidneys, eyes, sweat glands, and other body parts. Bacteria, plants, and invertebrates have water channels, too. But until Agre’s discovery, no one had isolated the molecules that form the channels. Ion channels help regulate the electrostatic potential of cells and mediate the transmission of nerve signals. MacKinnon was the first to determine an ion channel’s atomic structure and to figure out how the channel works.
Cells need specialized channels to import and export ions because ions are repelled by the fatty, nonpolar interior of the cell membrane. When Alan Hodgkin and Andrew Huxley demonstrated in the 1950s that nerve impulses consist of the fast, coordinated inflow and outflow of sodium and potassium ions, scientists knew that specialized portals for the two kinds of ion must exist in the membrane. The challenge was to identify the portals and explain how they worked. One functional aspect was particularly puzzling: How could a channel that admits K+ ions bar the smaller, identically charged Na + ions?
Nature’s need for water channels is less clear. Despite their polarity, water molecules are small enough to diffuse through the nonpolar interior of cell membranes with relative ease. But in certain parts of the body—notably the kidneys, which filter nearly 200 liters of water a day—diffusion is too slow. In 1970, Robert Macey demonstrated that exposing red blood cells to mercury ions impedes the flow of water across the cell membrane. Mercury ions stick to cysteine, an amino acid, so Macey’s experiment strongly suggested that a protein pore was involved in water transport. The challenge was to find the protein.
Aquaporins
Agre’s Nobel-winning work began in 1988—although not with the goal of finding water channels. At that time, he and his group at Johns Hopkins were trying to purify rhesus factor antigens taken from the membranes of red blood cells. Among the molecular leftovers was a 28-kilodalton protein of unknown origin and surprising abundance. Previous investigators had overlooked the protein because it doesn’t take up any of the usual histological stains.
To get an idea of the protein’s role, Agre ran its amino acid sequence through BLAST, a software tool that looks for genetic similarities. What BLAST revealed was intriguing. The 28-kDa protein’s genetic cousins are found in human kidneys, plant roots, cow eyes, and fly brains. Agre related the bizarre list to his friend and mentor John Parker, who realized what the members have in common: They process or contain water. Parker suggested that the unknown protein could be the elusive water channel.
To test the hypothesis, Agre adopted a technique used for studying ion channels. Channel-encoding RNA is inserted into frog egg cells, which have few membrane channels of their own. After a few days, the egg cells sprout genetically engineered channels.
Gregory Preston, a postdoc in Agre’s lab, prepared two sets of egg cells: one set equipped with the mystery protein and a control set without it. If the protein were a water channel, reasoned Agre, then placing a genetically modified egg cell in water would cause the cell to distend as osmotic pressure pushed water through the channels and into the cell.
The moment of discovery came on 9 October 1991. Preston dropped one of the genetically modified cells into distilled water and watched it through a microscope. In minutes, before he could position a video camera to record what was happening, the cell swelled, then burst. Next, he dropped a control cell into the water. It didn’t swell.
The group’s findings appeared in Science in 1992. 1 But despite the evidence, skeptics wanted to see the protein purified under physiological conditions and put to work as a water channel. Agre turned to the University of Pittsburgh’s Mark Zeidel, who incorporated the protein in liposomes and then measured the permeability using stopped-flow fluorescence. In that technique, a steady flow is abruptly terminated and the retreating front of the flow measured optically. Zeidel found that the protein indeed acts as a water channel and that it conducts water 10 to 100 times faster than a channel-free membrane. 2
Zeidel also found that the water channel is impermeable to a range of molecules and ions, including urea, a smallish molecule produced in the kidney, and hydrated protons (H3O+), which would upset a cell’s pH if they could get through. In view of its impressive selectivity, Agre dubbed the channel aquaporin.
Even before aquaporin had been identified as a water channel, Agre had noticed that a key sequence of amino acids, asparagine-proline-alanine (NPA, for short), cropped up in all the genetic cousins. He could also tell that the protein was made up of four units, each containing two NPA groups. By selectively mutating various parts of the protein, Agre’s group came up with a topological model in which the two NPA groups combined to form a narrow, ringlike portal.
Membrane-spanning proteins form three-dimensional crystals with notorious and frustrating difficulty. Though no cakewalk, it’s easier to coax the proteins to form two-dimensional crystals, which can then be subjected to electron microscopy. Using his helium-cooled electron microscope, Agre’s collaborator Yoshinori Fujiyoshi of the University of Kyoto determined the structure.
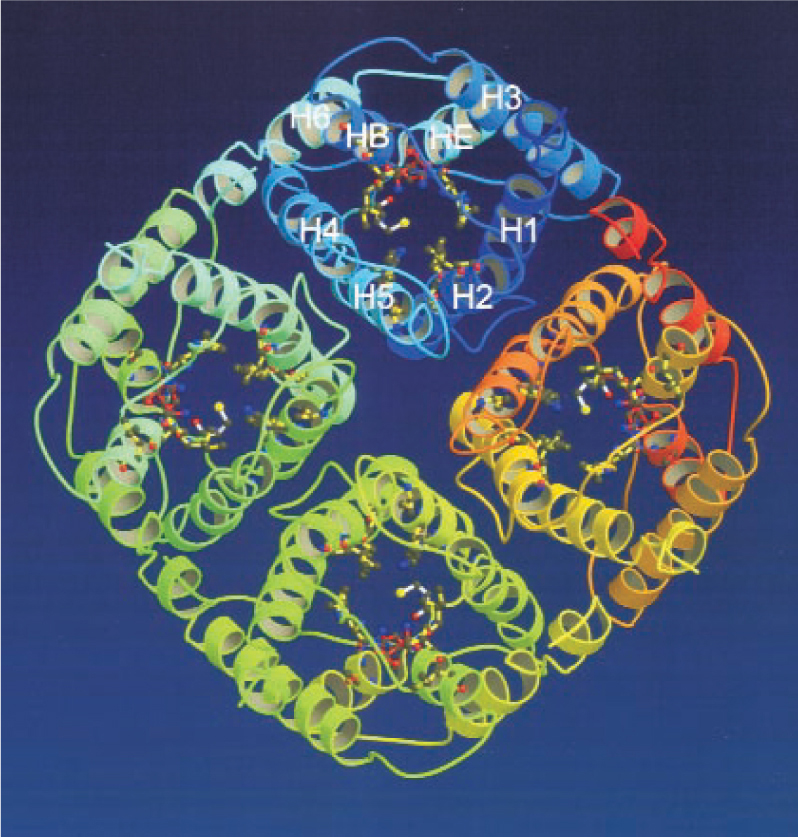
As figure 1 shows, the structure resembles a square cake cut into equal square quarters that are pulled apart to form a narrow vertical channel right down the center. At 3.0 Å at its narrowest point, the channel is a mere 0.2 Å wider than a water molecule.

Figure 1. Aquaporin-1, in ribbon representation, looking down the central channel. The eight alpha helices that make up each of the tetramer’s four subunits are designated H1–H8.
(Courtesy of Yoshinori Fujiyoshi.)
Such a narrow opening explains why larger molecules can’t go through, but what about protons? Water molecules flow down the narrow channel, oxygen first. With such an alignment, it’s easy for an oxygen atom at the top of the channel to form a hydrogen bond with a free proton and then give up one of its own protons to the next oxygen atom in line, and so on down the channel.
Aquaporin’s structure stops this proton-passing bucket brigade. As a water molecule arrives at the center of the pore, two asparagine residues lay hold to the molecule via hydrogen bonds and rotate it so that hydrogen atoms point up and down the channel. Thus the two hydrogen atoms of the water molecule can’t form hydrogen bonds with any adjacent water molecules and the transfer of protons is stymied.
Since the discovery of the first aquaporin (now called aquaporin-1), Agre and others have found about a dozen more in humans. Some have medical implications. Defects in aquaporin-2, for example, cause a rare but severe form of diabetes, while aquaporin-4, which is found in the brain, could play a role in dropsy. Says Agre, “the aquaporin proteins we’ve discovered and characterized are the answer to many of these physiological problems and will probably provide approaches to a number of disease states.”
Ironically, given their role in the aquaporin story, no one knows yet why red blood cells need water channels.
Ion channels
MacKinnon’s Nobel-winning work concerns ion channels and nerve cells. Inside a neuron, the concentration of K+ ions is about 30 times higher than outside. For Na + ions, the opposite is true. Large anions balance the charge inside the cell, while chlorine ions balance the charge outside the cell.
When a neuron is not transmitting a signal, the voltage across its membrane (measuring from out to in) is about −50 mV, thanks to the small concentration of K+ ions that seep out of the cell through so-called potassium leak channels. Further outflow of K+ ions down their steep concentration gradient is checked by the membrane’s negative potential. Sodium ions have both a concentration gradient and an electrostatic potential trying to push them into the cell. Yet somehow, despite their size (0.95 Å versus potassium’s 1.33 Å) they can’t go through the K+ leak channels.
Another kind of K+ channel operates during nerve transmission. When triggered by a voltage stimulus, Na + channels open to let Na + ions rush into the cell. The influx of positive charge drives up the membrane potential to +40 mV and triggers the opening of adjacent Na + channels ahead. Behind the pulse, however, the recently opened Na + channels enter a brief state of inactive closure during which they can’t respond to their usual voltage trigger. Because of this upstream inhibition, the pulse of positive voltage—the nerve signal—propagates forward along the neuron, but not backward.
Meanwhile, while the upstream Na + channels are locked shut, voltage-gated K+ channels open in response to the positive membrane potential. K+ ions rush out of the cell and quickly restore the membrane’s negative resting potential. This mechanism wouldn’t work if Na + ions could pass through the voltage-gated K+ channels.
The picture outlined above was established largely through electrophysiological experiments that measure the flow of various ions through membranes. Such experiments are exquisitely sensitive. For example, in 1955, Hodgkin and Richard Keynes proved that K+ ions cross the membrane two to three at a time.
The electrophysiological approach was complemented in the 1970s by genetic techniques. As an undergraduate at Brandeis University, MacKinnon worked with Chris Miller, an expert on K+ channels. When MacKinnon returned to Brandeis in 1985 as a postdoc, he joined Miller in preparing genetic variants of K+ channels and implanting them in frog egg cells. By comparing the ion channeling ability of the variants, MacKinnon determined that the K+ channel is a tetramer.
Other structural clues emerged as more channels were discovered. In organisms ranging from bacteria to humans, K+ channels of various types were found to have the same sequence of amino acids in common. In 1992, MacKinnon, then at Harvard University, showed that changing the shared sequence deprived K+ channels of their ability to obstruct Na + ions.
Despite this success, MacKinnon realized he wouldn’t be able to figure out how channels worked without seeing their three-dimensional structure. X-ray crystallography seemed the most promising approach, but to apply it, he had to do what no one had done before: create a sharply diffracting crystal of an ion channel protein.
Giving up his Harvard professorship, he moved to Rockefeller in 1996 and taught himself crystallography. Membrane proteins are hard to crystallize because their tops and tails are hydrophilic, while their middles are hydrophobic. The usual trick is to dissolve the membrane and its proteins with detergents. With the right detergent, protein–detergent complexes form crystals in aqueous solution.
Picking a detergent wasn’t a problem. Indeed, other labs had already crystallized ion channels. But those crystals, including MacKinnon’s first batches, made blurry, unusable diffraction patterns. MacKinnon thought the source of the blur might be the protein’s floppy ends, which he knew from his biophysical studies are not essential to channel function. The hunch paid off. He and his group used enzymes to nibble away the protein’s disorderly parts and were rewarded with clearly diffracting crystals. The effort took two years.
For the structure determination, MacKinnon took his crystals to the National Synchrotron Light Source at Brookhaven National Laboratory and to the Cornell High Energy Synchrotron Source. The 3.2 Å structure his group obtained appeared in Science in 1998 to immediate acclaim. 3
The paper and its more detailed successor 4 outlined how the channel works. In aqueous solution, K+ and Na + ions surround themselves with water molecules whose oxygen atoms are attracted by the ions’ positive charges. To draw ions through its narrow entrance, the channel has to remove the water molecules. It can do this, thanks to rings of carbonyl (C=O) groups that line the entrance. Held in place by a rigid cuff, the carbonyl groups have just the right configuration to take over the role of stabilizing K+ ions from the water molecules.
Sodium ions, however, are too small to feel the combined influence of the carbonyl groups. They remain hydrated and outside the cell.
A single K+ ion at the head of the channel would stay put, but, as figure 2 shows, the entrance is long enough to accommodate more than one ion at a time. Mutual electrostatic repulsion ensures that the ions keep moving, but to cross the membrane, they must pass through the membrane’s nonpolar interior.

Figure 2. The narrow entrance of a K+ channel has four sites where negatively charged oxygen atoms stabilize K+ ions (green). The ions′ mutual electrostatic repulsion pushes them through the entrance and down into the water-filled cavity, where water molecules provide electrostatic stability.
(Courtesy of Roderick MacKinnon.)
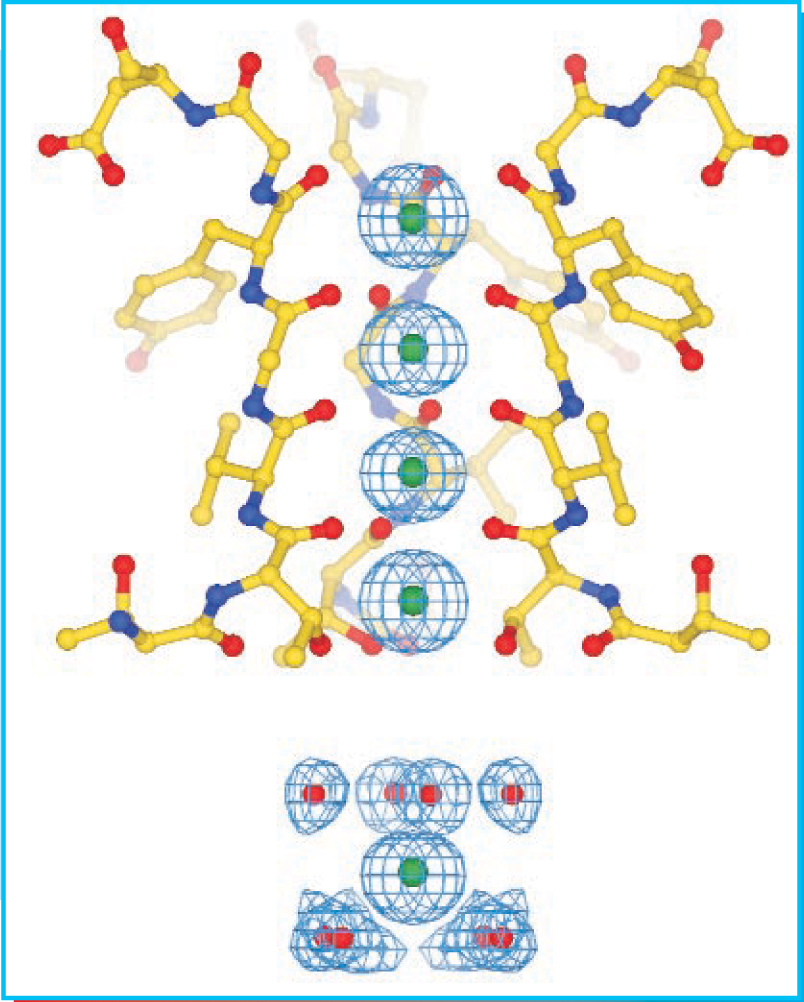
The narrow entrance opens into a water-filled cavity at the center of the channel. Once inside the cavity, an ion attracts water molecules, but their stabilizing effect isn’t strong enough to mitigate the electrostatically unwelcoming environment of the nonpolar interior. MacKinnon’s structure shows that long-range electrostatic help comes in the form of negatively charged substructures (alpha-helices) that impinge on the cavity’s lining. In an aqueous environment, dissolved ions screen out such long-range interactions, but in the nonpolar environment of the membrane, the stabilizing power of the helices is undiminished.
Since his pioneering work, MacKinnon and his collaborators have continued to work on K+ channels and, recently, on chlorine channels. Like Agre, he praises his many young collaborators. “One of the great things about being a scientist in this day and age,” he says, “is that talented people come from all over the world to work with you. Without that, none of this would ever happen.”
Biographies
Peter Agre was born in 1949 in Northfield, Minnesota. He earned a BA in chemistry from Augsburg College in 1970 and an MD from Johns Hopkins in 1974. After postdoctoral work at the University of North Carolina, he returned to Johns Hopkins, where he has remained ever since.
Roderick MacKinnon was born in 1956 in the Boston suburb of Burlington. He earned a BA in biochemistry from Brandeis in 1978 and an MD from Tufts University in 1982. After postdoctoral work at Brandeis, he joined the faculty of Harvard Medical School in 1989. He moved to Rockefeller, his current academic home, in 1996.

Agre
JOHNS HOPKINS UNIVERSITY


MacKinnon
ROCKEFELLER UNIVERSITY

References
1. G. M. Preston, T. P. Carroll, W. B. Guggino, P. Agre, Science 256, 385 (1992) https://doi.org/10.1126/science.256.5055.385 .
2. M. L. Zeidel, S. V. Ambudkar, B. L. Smith, P. Agre, Biochemistry 31, 7436 (1992) https://doi.org/10.1021/bi00148a002 .
3. D. A. Doyle, J. M. Cabral, R. A. Pfuetzner, A. Kuo, J. M. Gulbis, S. L. Cohen, B. T. Chait, R. MacKinnon, Science 280, 76 (1998) https://doi.org/10.1126/science.280.5360.69 .
4. Y. Zhou, J. H. Morais-Cabral, A. Kaufman, R. MacKinnon, Nature 414, 43 (2001) https://doi.org/10.1038/35102009 .





{kind=link}
{kind=link}
{kind=link}
{kind=link}