Membrane phase demixing seen in living cells
DOI: 10.1063/PT.3.3838
To keep their insides in and their outsides out, cells and their substructures are enveloped by lipid bilayer membranes. The membranes have the consistency not of rubbery balloons but of thin liquid films: Proteins and other molecules embedded in the membranes can diffuse from place to place. But despite that diffusion, the membranes are compositionally heterogeneous, not uniformly mixed, and they sometimes appear to host discrete domains enriched in particular molecules.
Biologists and biophysicists have long debated the mechanism of membrane organization—and, in particular, whether it has anything to do with thermodynamic phase separation, the same physics that drives the demixing of a shaken bottle of vinegar and oil. Now Sarah Keller, Alexey Merz, and their colleagues at the University of Washington in Seattle have shown that membrane domains do indeed form through phase separation, at least in some living cells. 1
Keller’s interest in the problem was from a physics point of view: How can cell membrane organization be understood in terms of what’s known about phase separation in other liquids? Merz came to the topic from a cell biology perspective: How might membrane structure relate to the formation and function of cells and organelles? Together, they co-mentored the brother–sister team of graduate students Scott Rayermann (pictured in figure
Figure 1.

Scott Rayermann prepares yeast vacuole membranes for fluorescent imaging.
GLENNIS RAYERMANN

The Seattle researchers looked at the membranes that enclose vacuoles—a type of organelle—in single-celled budding yeast. They sought and found two telltale hallmarks of phase-separation dynamics. First, when two domains with the same composition coalesced, the surrounding domain boundaries quickly rearranged to minimize the overall energy due to line tension. Second, when the yeast cells were heated and cooled, the domains disappeared and re-formed—and the transition consistently happened at a single characteristic mixing temperature.
Vesicle models
One reason membrane organization has been so difficult to study is that the putative domains are so small. Experiments to date on the membranes that surround whole cells tend to rule out domains larger than tens of nanometers. That’s too small to see with conventional optical microscopy. Superresolution techniques can help to bring tinier structures into view, but the gain in resolution is accompanied by a loss of speed, so crucial information about dynamics is lost.
Cell membranes, however, are not free-standing bubbles. They are anchored to the cytoskeleton inside, and the anchoring points may be preventing domains from growing any larger than the distance between attachments.
For a simpler system, researchers have turned to synthetic membranes. Under some conditions, lipids in water spontaneously assemble into bubble-like membranes called vesicles. And with some coaxing, vesicles can be grown to sizes of 100 µm or more, so they and their correspondingly larger domains can be optically imaged with ease. It’s been observed since the early 2000s that vesicles made from a combination of lipids—one of the simplest formulations is two lipids with different melting points plus cholesterol—exhibit complex phase behavior, with the components mixing at high temperatures and segregating into immiscible domains when cooled.
2
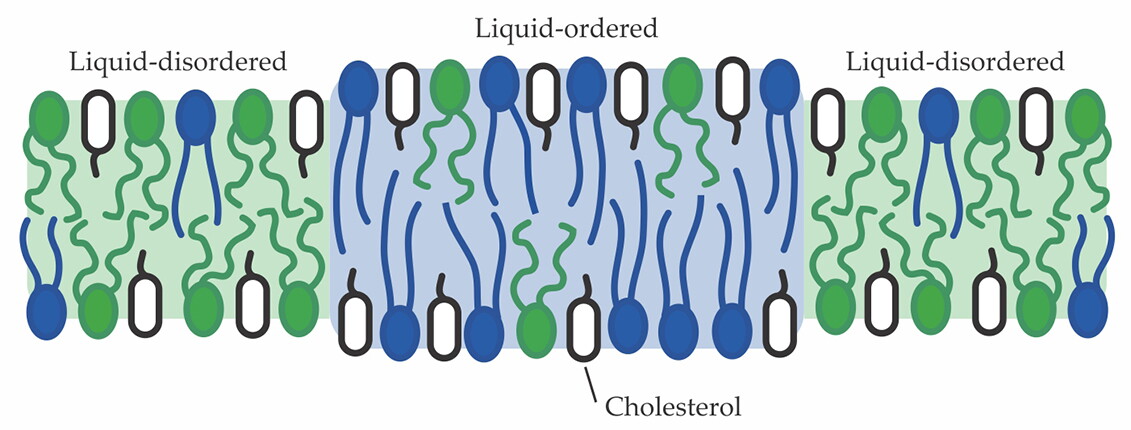
Those compositionally distinct ordered and disordered phases, shown schematically in figure
Figure 2.

In a model synthetic membrane made of two types of lipid (green and blue) and cholesterol (white), the components separate into immiscible liquid-ordered and liquid-disordered phases below a characteristic mixing temperature. In biological membranes, which contain proteins embedded in the lipid layers, those molecules may be preferentially partitioned into one phase or the other. (Courtesy of Sarah Keller.)
The synthetic vesicles were an interesting model system that yielded new physics discoveries. 3 But the question remained of their relevance to biology. Living cell membranes contain not just lipids and sterols but a multitude of embedded proteins. Certain biological membranes showed phase-demixing behavior similar to that seen in vesicles, but only when they’d been removed from their cellular environment or otherwise perturbed from their physiological state. And they had some curious properties: Membranes taken from a culture of zebrafish cells, for example, consistently underwent phase demixing at temperatures 17 °C below the cell-growth temperature—even when the cell-growth temperature was changed. 4 But observing a demixing transition in vivo, or even under physiological conditions, remained frustratingly elusive.
A turning point came in 2013. Alexandre Toulmay and William Prinz, of the National Institutes of Health in Bethesda, Maryland, noticed that when yeast cells entered the stationary growth phase—when a dearth of nutrients prompted the cells to temporarily give up on trying to reproduce—their vacuole membranes formed micron-sized domains strikingly similar to the liquid-ordered and liquid-disordered phases in synthetic vesicles. 5 Moreover, when the researchers fluorescently labeled 14 different membrane proteins, they found that each one was enriched in one of the two membrane regions and depleted in the other.
Vacuole phases
Just because the vacuole domains looked like liquid-ordered and liquid-disordered phases, however, doesn’t necessarily mean that they were. Although Toulmay and Prinz tried heating and cooling the cells, they saw no signs of a mixing transition.
The Seattle researchers were inspired to pick up where the NIH team left off: If a phase-separation mechanism was responsible for the vacuole domains, they were determined to see dynamic proof of it. “Two of our assets are stubbornness and optimism,” says Keller. “According to Toulmay and Prinz’s paper, our experiments should not have worked. We did them anyway.”
The biggest challenge was in imaging the tiny structures. The vacuole domains were bigger than the diffraction limit of conventional microscopy, but seeing them clearly still required sophisticated optical tricks.
Vacuoles are spherical and transparent. Typically, a reconstructed projection of a 3D object would comprise superimposed images of the front and back hemispheres, which could have obscured important features. So the researchers used deconvolution microscopy to obtain stacks of 2D slices, from which they painstakingly reconstructed images of just one hemisphere.
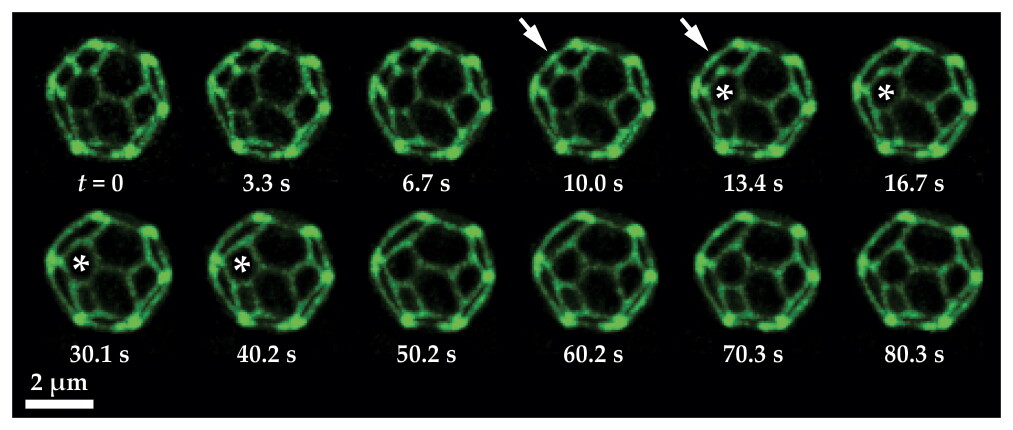
Figure
Figure 3.

Domain coalescence on short time scales in this series of optical micrographs demonstrates that the domains are liquid. The green lines mark the distribution of a fluorescently labelled protein that’s partitioned into one of the two membrane phases; the green phase forms thin boundaries between domains of the other phase. When the boundary indicated by the arrows breaks, the surrounding domains quickly rearrange. In particular, the domain marked by the star changes from hexagonal to pentagonal. (Adapted from ref.
To observe the other manifestation of liquid phase separation—the mixing and demixing of domains—the researchers turned to a version of light-sheet microscopy 6 that conferred the necessary combination of low noise and high imaging speed. Above the transition temperature of about 38 °C, the domains mixed and the vacuoles fluoresced a uniform pale green. Below the transition temperature, dark domains on a green background reemerged.
Useful transitions
The results confirm that vesicle models are indeed relevant to at least some biological systems, but it remains to be seen whether membrane domains in other organelles and species form by the same mechanism. “Our work implies that all biological membranes have the opportunity to employ miscibility phase transitions,” says Keller. “However, there’s no obligation for them to do so.”
What does it mean for a cell to employ a phase transition? If each membrane protein is confined to either the liquid-ordered or the liquid-disordered domains, domain formation brings some proteins closer together and keeps others farther apart. If a cellular process is helped or hindered by contacts between particular membrane proteins, then the demixing transition could serve as a way to switch the process on or off in response to changes in temperature. In fact, recent work by Jodi Nunnari and collaborators suggests that a critical cell-growth pathway in budding yeast may be regulated in just that way. 7
In nature, yeast cells encounter a wide variety of environmental conditions, and they may sometimes benefit from switching at temperatures other than 35–40 °C. Keller, Merz, and colleagues are planning to investigate whether and how the transition changes in different yeast strains to help them adapt to different environments.
References
1. S. P. Rayermann et al., Biophys. J. 113, 2425 (2017). https://doi.org/10.1016/j.bpj.2017.09.029
2. C. Dietrich et al., Biophys. J. 80, 1417 (2001); https://doi.org/10.1016/S0006-3495(01)76114-0
S. L. Veatch, S. L. Keller, Biochim. Biophys. Acta 1746, 172 (2005). https://doi.org/10.1016/j.bbamcr.2005.06.0103. See, for example, A. R. Honerkamp-Smith, B. B. Machta, S. L. Keller, Phys. Rev. Lett. 108, 265702 (2012). https://doi.org/10.1103/PhysRevLett.108.265702
4. M. Burns et al., Biophys. J. 113, 1212 (2017). https://doi.org/10.1016/j.bpj.2017.04.052
5. A. Toulmay, W. A. Prinz, J. Cell Biol. 202, 35 (2013). https://doi.org/10.1083/jcb.201301039
6. M. Tokunaga, N. Imamoto, K. Sakata-Sogawa, Nat. Meth. 5, 159 (2008). https://doi.org/10.1038/nmeth1171
7. A. Murley et al., J. Cell Biol. 216, 2679 (2017). https://doi.org/10.1083/jcb.201610032
More about the authors
Johanna L. Miller, jmiller@aip.org





{kind=link}
{kind=link}
{kind=link}