The emergent physics of animal locomotion
DOI: 10.1063/PT.3.3691

Moths flitting between flowers on a moonlit night and cockroaches scurrying underfoot are dynamical systems. Like many other animals, they get around with a seeming ease and agility that we humans find hard to replicate in systems we create. It may seem that we ought to know everything there is to know about animal locomotion. But we have yet to meet Richard Feynman’s provocative standard, “What I cannot create, I do not understand.” The failure is not due to a limitation of our engineering abilities; rather, it reflects the difficulty of puzzling out how movement emerges from the physical and physiological systems of organisms. We cannot yet emulate the motility seen in nature nor derive the behavior.
The mystery is not new. In his prescient What Is Life? (1944), Erwin Schrödinger wrote about the possible limits of reductionism in explicating the physics of life and recognized that new principles might arise from examining life at the organism scale:
From all we have learnt about the structure of living matter, we must be prepared to find it working in a manner that cannot be reduced to the ordinary laws of physics. And that not on the ground that there is any “new force” or what not, directing the behaviour of the single atoms within a living organism, but because the construction is different from anything we have yet tested in the physical laboratory.
Today a physics approach to complex systems and a biomechanics approach to living systems are being integrated in the physics laboratory.
Neuromechanics, a science of movement
All animals can move. They (and some plants, fungi, and prokaryotes) have evolved diverse movement strategies that rely on generating stable and maneuverable dynamics even when the world is uncertain, slippery, compliant, or flowing.
When moving, animals must acquire, process, and act on information from neurons, muscles, the body, and the external environment. Figure
Figure 1.

Animal movement, as represented by a neuromechanical system diagram. Whether in humans or moths, neurons activate muscles by transmitting electrical impulses over time (t)—frequently digital, all-or-nothing events termed action potentials or spikes. Muscles transform the time-varying neural motor commands u(t) into mechanical forces f(t). Muscles actuate the body, but it is the interaction of the body with the surrounding environment that produces locomotion y(t). Meanwhile, myriad sensors detect not only the body and how it interacts with environment, but also internal states of the animal. All that information feeds back through the animal’s nervous system. Sensor signals can be combined with reference signals, illustrated by red arrows. In this schematic, r1(t) represents a brain-directed task such as “fly at a steady speed,” and r2(t) represents exogenous sensory cues such as a moth might receive when tracking the movement of a flower.
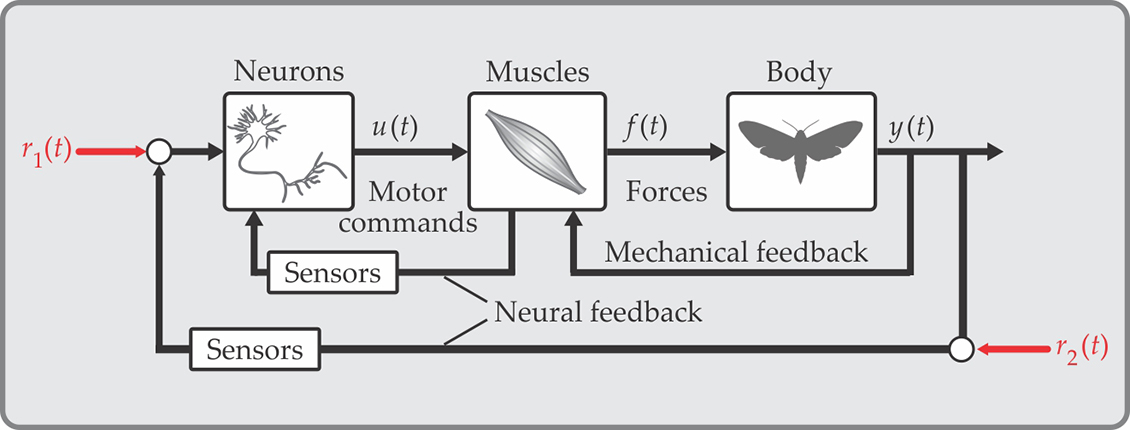
Neuromechanics considers how movement arises through the interplay of multiple physiological systems and their interactions with the environment around the animal. Understanding how those systems control movement remains elusive in part because no one animal can serve to test all neuromechanical hypotheses. Researchers can, however, turn to the idea that “for such a large number of problems there will be some animal of choice, or a few such animals, on which it can be most conveniently studied,” as August Krogh wrote in his 1929 American Journal of Physiology article, “The progress of physiology.”
To fulfill Schrödinger’s hope for new laws of organisms, we can discover themes in individual well-suited exemplars, as Krogh’s principle would encourage. But we also can systematically relate the dynamics of movement across many species and explicitly incorporate the shared evolutionary history of organisms. We can also hope, with experimental input, to determine general principles that span organisms and that could translate even to nonbiological systems such as robots.
The interface of physics and organismal biology has both precedence and promise. The field of neuromechanics is quite mature, and a comprehensive review would neither be fruitful nor serve to motivate the physicist who has not been exposed to the field. Rather, I’ll share a few examples, some of which motivated my own research. After exploring the insights provided by those examples, I’ll turn to emerging frontiers. Throughout this perspective, I will highlight discoveries and examples that come from studying the world of insects, a group of animals with incredible diversity and, in particular, a wide range of locomotive behaviors. Insects are not particularly simple, especially from a physics perspective, but they are relatively small, have short generation times, and are amenable to many established and emerging experimental tools.
Insect flight and unsteady aerodynamics
The images on page
Figure 2. Visualizing airflow. The airflow lines (a) over the wing of the twilight flying hawk moth (Manduca sexta) form bound leading-edge vortices (LEVs; blue curves). The force from an LEV, FLEV, enhances both the lift and the drag force on the wing. The orange line is a chord that connects the leading and trailing edges of the wing. (b) Curved airflows due to LEVs are evident in this photo (courtesy of Megan Matthews) of a moth flying through a plane of smoke in a wind tunnel.
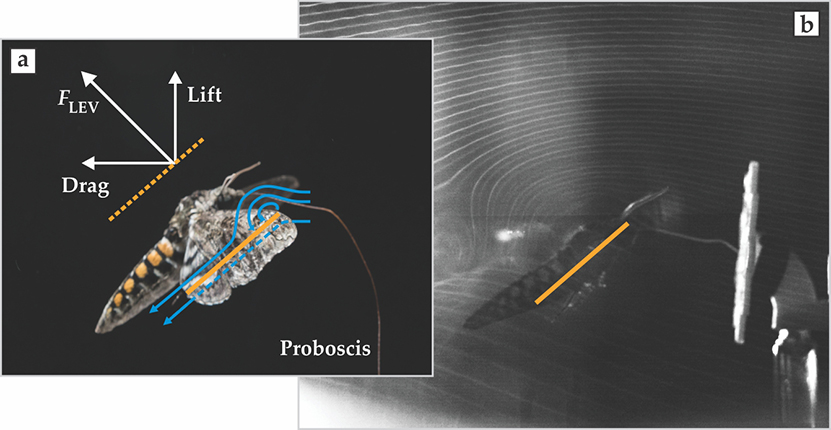
The story goes back to the 1970s when zoologist Torkel Weis-Fogh examined the chalcid wasp (Encarsia formosa) and discovered a new mechanism by which small animals with flapping wings generate the lift they need to hover and maneuver. 1 At the end of the wasp’s upstroke, the insect’s wings clap together before separating and flinging outward, a maneuver that allows air to refill the vacated space. It was applied mathematician James Lighthill who characterized the mechanics behind the clap-and-fling action. 2 The key idea is that the separation of two clapped wings creates equal and opposite circulations on each wing during the fling. Those trapped circulations produce added lift because the air flowing over them has to be directed downward toward the surface to reattach and keep the vortex in place. The redirection effectively sucks the wing normal to its surface and thus creates lift and drag. Lighthill noted that it was “surprising that a fundamentally new mechanism of lift generation in inviscid two-dimensional flow should be discovered six decades after the work of Prandtl, Zhukovski, Kutta and Lanchester.”
It turns out that most flapping animals do not clap their wings together fully, so the specific clap-and-fling behavior described by Weis-Fogh and Lighthill cannot generally explain insect flight. However, the seeds of future discovery were sown by the insights that flow remains bound to wings and that time-dependent aerodynamic mechanisms might be critical. Still, not until more than 20 years later, in the late 1990s and early 2000s, was a more general mechanism identified for how insects fly: As the wing moves, the flow around it rolls up to produce a bound vortex on its leading edge
3
as shown diagrammatically in figure
But LEVs are far from the end of the story. 4 Other unsteady aerodynamic mechanisms may enhance lift and modulate flight forces. During flight, a fly’s wing interacts with the vortex wake shed from the previous wing stroke. The animal may be able to recapture energy from those interactions, a process that would provide a fascinating illustration of how the environment can be used to store and return energy. Also, it remains unclear how LEVs persist or are modified when the animal encounters air that is already disrupted. Nearly all studies about the mechanisms of flight have been performed in still or steadily flowing air. If the moth and other flapping insects and birds rely on binding vortices to their wings, how do they produce, stabilize, and modulate those in environments where the fluid is already unsteady or even fully turbulent?
Simple models, important insights
Emergent behaviors are complex patterns that result from the interactions of components that are simple, or at least simply described, and that don’t display the emergent patterns of the whole. Understanding emergent behavior of physiological systems or whole organisms can be frustratingly difficult. After all, the system components themselves are complex, heterogeneous structures involving neurons, muscles, limbs, and the matter surrounding the animal. Yet in many cases, low-dimensional dynamics models—that is, models with just a few degrees of freedom—sometimes called templates, can describe gaits and movement strategies. 5
One such template model describes the way in which many animals run, bouncing up and down like a spring-loaded inverted pendulum.
6
Such SLIP dynamics, illustrated in figure
Figure 3.

Template models of running. Nearly all running animals use two sets of legs that alternately push off the ground. (a) For the cockroach, the two alternating sets are tripods, indicated by the gray and blue arrows. The vertical movement of the animal’s center of mass (black-and-white filled circles) looks like trajectories of a spring-loaded inverted pendulum (dashed curves) pieced together where one leg tripod lifts off and the next touches down. The jagged lines represent effective compressed springs exerting forces on the cockroach; the red spring indicates maximum compression. (b) Lateral movement can also be modeled as arising from spring forces. The line of force doesn’t always pass through the center of mass, so the cockroach is periodically torqued as it runs.
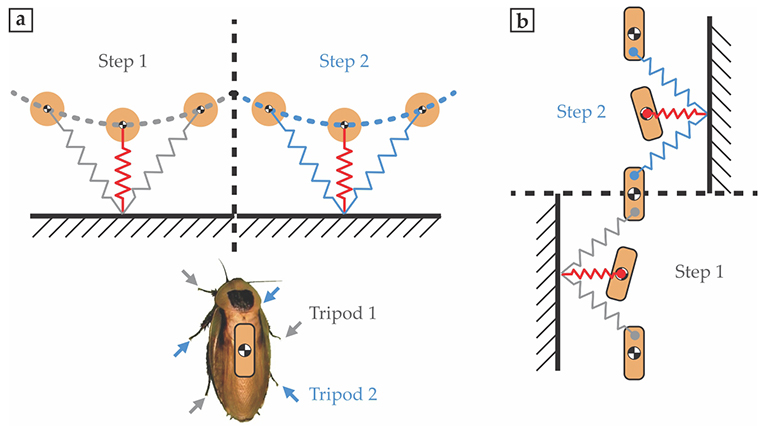
Dynamics in the lateral plane can be even simpler. As illustrated in figure
The SLIP and lateral dynamics conserve energy during each of the animal’s steps. An energy-conserving dynamic system should not be stable to perturbations—for example, a small change in external force due to a bump in terrain—because there is no mechanism for dissipating energy generated by means of those perturbations. Yet the simple lateral template can give stable motion. The reason can be traced to the discontinuity in the animal’s gait, when one limb or set of limbs picks up and the next set touches down. At that time, the direction of the spring force suddenly switches, a transition sufficient to stabilize the motion.
Apparently, animals, and robots too, can achieve dynamic stability just by maintaining their steady, alternating gait mechanics. My colleague Robert Full and I tested that notion by putting cockroaches on rough terrain and recording the neural commands they sent to their muscles. 8 The two of us found that many of the muscles in the cockroach leg did not change their activation, which suggests that the mechanical motion is indeed stable, at least until perturbations get so large that the cockroach must actively modify its behavior. Moreover, legged robots that simply maintain an alternating tripod gait can run stably over a suitably scaled version of the same rough terrain, even if they have no external sensors. The robots even achieve peak speed if their size-adjusted leg stiffness is similar to that observed in animals.
The simple template models discussed above are not a panacea for the complexity of living systems. They are a starting point. Their simplicity fails to capture many specifics of how animals move. Damping is critical in most biological movement, robust stability typically requires neural feedback in some measure, and control of movement depends on the interaction of limbs with their surrounding environment. Scientists are just beginning to understand the physics of what happens when moving, deformable bodies encounter a nonrigid terrain with high friction. Template models do, however, provide important insights that must be embedded into more complex models. The key is to understand the scales at which new dynamical features emerge.
Quantitative patterns of behavior
Many fewer physicists have tackled problems in macroscopic or even mesoscopic living systems than have contributed to understanding those systems at the molecular and, more recently, cellular level. In the past, that reluctance may have had much to do with cultural and technological limitations. One of the most significant constraints on linking physics to locomotion was the challenge of capturing complex movements with sufficient scope and precision to extract quantitative patterns. As high-speed imaging, machine vision (automated inspection), motion capture, and rapid prototyping have transformed how scientists understand movement, those limitations have started to fall away. It is now feasible to collect thousands of steps, wing strokes, and other motions in a single experiment.
Of course, quantification alone is not sufficient for a science of movement. Using approaches from statistical physics, biophysicists can identify relatively simple patterns and low-dimensional structures present in the movements of animals. 9 One recent decomposition of movements showed that a mere four degrees of freedom suffice to capture 95% of the movements of the small roundworm Caenorhabditis elegans—for example, two of the dimensions determine the phase of the oscillatory body wave. Low-dimensional reconstructions of cockroach gaits can separate the time courses of neural and mechanical stabilization responses to perturbations. Machine vision has shown that a surprisingly small set of probability density functions suffice to describe repetitive fruit-fly behaviors such as running or wing grooming and even to capture the differences among individual preferences.
In conjunction with quantitative studies of behavior, researchers can now electrically and optically modify neural and muscular signals to alter behavior. For example, optogenetics involves the expression of light-sensitive proteins that alter the electrical activity of individual nerve cells. 10 Gene silencing and activation tools let researchers selectively manipulate neurons, muscles, and sensors. It used to be that such tools were limited to a few model organisms, but the range of accessible animals is expanding thanks to rapid sequencing and genome editing tools such as the CRISPR/Cas-9 toolkit.
More techniques become available every year, and large initiatives are emphasizing the newly accessible data sets they produce. Perhaps largest of all is the BRAIN (Brain Research through Advancing Innovative Neurotechnologies) Initiative, 11 a multibillion-dollar investment by government agencies, universities, and private foundations that will run through 2025. Brains have an enormous number of neurons; even the cockroach brain has 105–106 neurons. The BRAIN Initiative will include efforts focused on developing new technologies and analytical tools and creating atlases of neurons and their connections. The hope is to meaningfully address the question “How does the brain work?” It’s a wonderfully inspiring question, but one that is difficult to answer in view of the many contexts in which brains might operate. How can a physics perspective help scientists understand how complex living systems behave, given the proliferation of methods to interrogate them?
What is needed is a way to move from the large volumes of data provided by our new tools to principles of movement. Three approaches are worth highlighting because they have already had some success and are providing a constructive path forward: experimental manipulation of feedback dynamics, multiscale analysis of living assemblages, and robotic modeling.
Altering feedback dynamics
Ever since Norbert Wiener’s 1948 book Cybernetics, or Control and Communication in the Animal and the Machine, the concept of feedback has been tied to the behavior of many dynamic systems, living and engineered. Feedback occurs when the output of a dynamic process changes the input into that process. It is not usually emphasized in a physics approach to dynamics, but its ubiquity in living systems means it cannot be ignored. Moreover, introducing even simple feedback into a system can have profound effects on both the qualitative and quantitative dynamics of movement. 12
Figure
Over the past few years, neuromechanists have begun systematically to explore function and mechanism by changing the structure or context of feedback. An example I was involved with
13
featured the twilight flying hawk moth shown on page
To test the idea, my colleagues and I observed moths tracking robotic flowers at different light levels. By using simple feedback dynamics, we could factor out the moth mechanics and isolate the change in neural processing due to a drop in light intensity. We found that moths do in fact slow their processing to increase sensitivity, but only to the extent that they can still accurately track flower movements below about 2 Hz. That seemed odd until we took videos of moth-pollinated flowers blowing in windy environments and found that nearly all flower movements occur at frequencies just below that threshold. The moths, it seems, match their neuromechanics to the dynamics of their preferred flowers.
Multiscale physics of living assemblages
Brains, muscles, and other physiological systems are organized into multiscale networks. That network structure can lead to unexpected emergent dynamics, especially when network subunits are active—that is, capable of producing forces on their environment. Experiments, simulations, and analyses that connect processes at multiple scales are cutting through the difficulty of understanding aggregations, even of heterogeneous living components.
Consider, as a specific example, the structure of muscle, nature’s most versatile actuator. As shown in figure
Figure 4.

Muscle as an active crystal. Muscle is a hierarchical structure with an organization spanning at least seven orders of magnitude in size, from 10−8 m to 10−1 m. (a) Muscle structures include fibers, subcellular structures known as sarcomeres, and, at the smallest scale, a lattice of the filamentous proteins myosin (red) and actin (blue). Myosin has an active motor region that reaches across to actin filaments and then binds and ratchets the actin so that it slides relative to the myosin filament. (Whole-muscle diagram adapted from a National Institutes of Health image. Filamentary-lattice image courtesy of Dave Williams.) (b) The billions of myosin motors arranged along the actin filaments make the protein lattice active—that is, capable of producing internal stress and thus changing the lattice spacing. The crystalline protein lattice diffracts x-ray light and produces a pattern that depends on the spacing and arrangement of the protein filaments. (Image courtesy of Travis Tune.)
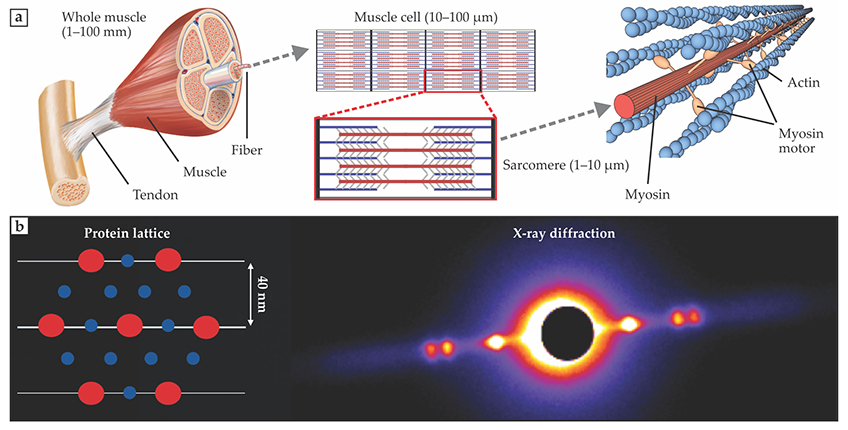
A muscle can include billions of individual molecular motors that grab onto and constrict the filamentary lattice or put the filaments under tension. From x-ray imaging, we have learned that a mere nanometer change in lattice spacing can dramatically change the macroscopic behavior of muscle. For example, cooling the temperature of the moth’s main flight muscles by as little as 10 °C can lock up the lattice and cause a muscle that acts like a motor when warm to store and return energy like a spring when cool. With the help of 3D simulations, biophysicists are beginning to connect the macroscopic behavior of a muscle to the collective mechanics of individual molecular motors organized in a hierarchical structure. 15
Collective action is also important outside the confines of a single living organism, particularly when organisms behave in aggregations. Flocking and schooling are perhaps the best-known examples of such collective behaviors, and they have interesting energetic and ecological consequences. Other collections of active biological agents manifest a diverse range of emergent properties and behaviors. Some aggregations allow organisms to adopt remarkable material properties and manipulate their environment. 16 In the insect world, colonies of ants form rafts and bridges that allow them to span gaps and survive floods. The collective action of honeybees can control the fluid dynamics of the hive and thus aid in the ventilation of their home.
Robophysics
Robots are becoming ubiquitous and are even finding their way into the physics laboratory. Robophysics is the use of robotic physical models to examine a principle of movement or control without having to deal with the complexity and experimental constraints of living organisms. Robots have already transformed the study of organismal biology and are now helping physicists make sense of active, motile systems—living, as in the examples shown in the top panels of figure
Figure 5.

Robot analogues. When running on rough terrain, (a) the cockroach Blaberus discoidalis uses stable mechanical dynamics requiring minimal feedback control. (b) The same dynamics are realized in the robotic hexapod RHex when it moves over a version of the same terrain. (Adapted from U. Saranli, M. Buehler, D. E. Koditschek, Int. J. Robot. Res. 20, 616, 2001, doi:10.1177/02783640122067570 .) (c) The subsurface swimming sandfish lizard (image courtesy of Rob Felt) inspired (d) a robotic model that captures the undulatory pattern used by the lizard to move through granular material (image courtesy of Daniel Goldman). (e) The vinegar fly Drosophila suzukii (image courtesy of Martin Cooper) and (f) fly-inspired robots (adapted from K. Y. Ma et al., Science 340, 603, 2013, doi:10.1126/science.1231806 ) use flexible wings, specialized actuators, and a deformable exoskeleton to flap at frequencies above 100 Hz.

Early on, robotic models revealed new biology and new physical mechanisms in studies of animal movement in fluids. 18 Aquatic swimmers were some of the first systems to be explored from a robotic modeling perspective. James Gray’s 1936 attempt to reconcile the power of dolphin locomotion with the high drag of hydrodynamic models sparked many decades of examination into drag reduction in turbulent boundary layers. Ultimately, though, the paradox he raised was resolved with improved imaging and a better understanding of muscle power production. Nonetheless, discoveries about animal hydrodynamics have inspired several generations of fish robots and even new wind turbine designs based on the flukes of the humpback whale.
More recently, the role of LEVs on insect wings was rigorously explored through the use of a robotic model of fly wings scaled up and dipped in mineral oil to maintain a Reynolds number comparable to that of living flies. And just within the past couple of years, controlled flight and stable hovering have been engineered in a centimeter-sized aerial robot with a mass of 0.01 g.
Robophysical models have become an important complement to theory and conventional simulations and experiments. They have even brought forth a new field, terradynamics, which addresses how motile systems interact with a complex granular medium such as sand, snow, or mud. Studies have revealed how snakes sidewind, how sandfish swim through sand, and how early legged animals might have first crawled up on muddy shores. As a consequence of the deeper understandings they have provided, robotic models have often inspired engineering advances as well.

The twilight flying hawk moth. Manduca sexta hovers and feeds from a robotic flower in a laboratory flight chamber. (Photo by Rob Felt, Georgia Tech.)

What really marks the maturation of robophysics is the proliferation of tools that lower the bar for the creation of robotic systems; those include rapid prototyping, 3D and 4D printing, “smart” materials, and cheap, easy-to-learn commercial component systems. A student of physics or biology can now rapidly build a functioning robotic platform to test how compliant legs interact with yielding ground or how flexible wings deform during flapping. We can now create in order to understand.
An opportunity
A physics perspective on neuromechanics looks to extract principles from the dynamic and information-processing properties of moving organisms. For the physicist interested in the workings of nature’s most versatile systems, neuromechanics represents an exciting opportunity. Scientists’ understanding of such macroscopic phenomena as the agile movement of the hawk moth remains incomplete, but we can expect to glean new principles if we can cut through the enormous swaths of data afforded by observing living systems with modern tools. If we can avoid simply reveling in the complexity of the biological world and instead work to understand the relevant features of complex systems, we will find ample opportunity to apply a physics perspective to the study of movement and behavior, even at the level of the organism.
References
1. T. Weis-Fogh, J. Exp. Biol. 59, 169 (1973).
2. J. M. Lighthill, J. Fluid Mech. 60, 1 (1973). https://doi.org/10.1017/S0022112073000017
3. C. P. Ellington et al., Nature 384, 626 (1996). https://doi.org/10.1038/384626a0
4. For reviews, see S. P. Sane, J. Exp. Biol. 206, 4191 (2003); https://doi.org/10.1242/jeb.00663
D. Lentink, Bioinspir. Biomim. 9, 020301 (2014); https://doi.org/10.1088/1748-3182/9/2/020301
Z. J. Wang, Annu. Rev. Condens. Matter Phys. 7, 281 (2016). https://doi.org/10.1146/annurev-conmatphys-031113-1338535. R. J. Full, D. E. Koditschek, J. Exp. Biol. 202, 3325 (1999);
P. Holmes et al., SIAM Rev. 48, 207 (2006). https://doi.org/10.1137/S00361445044451336. R. Blickhan, R. J. Full, J. Comp. Physiol. A 173, 509 (1993).
7. J. Schmitt et al., Biol. Cybern. 86, 343 (2002). https://doi.org/10.1007/s00422-001-0300-3
8. S. Sponberg, R. J. Full, J. Exp. Biol. 211, 433 (2008). https://doi.org/10.1242/jeb.012385
9. G. J. Stephens et al., PLOS Comput. Biol. 4, e1000028 (2008); https://doi.org/10.1371/journal.pcbi.1000028
S. Revzen et al., Biol. Cybern. 107, 179 (2013); https://doi.org/10.1007/s00422-012-0545-z
G. J. Berman et al., J. R. Soc. Interface 11, 20140672 (2014). https://doi.org/10.1098/rsif.2014.067210. K. Deisseroth, Nat. Methods 8, 26 (2011). https://doi.org/10.1038/nmeth.f.324
11. T. R. Insel, S. C. Landis, F. S. Collins, Science 340, 687 (2013). https://doi.org/10.1126/science.1239276
12. K. J. Åström, R. M. Murray, Feedback Systems: An Introduction for Scientists and Engineers, Princeton U. Press (2008);
E. Roth, S. Sponberg, N. J. Cowan, Curr. Opin. Neurobiol. 25, 54 (2014). https://doi.org/10.1016/j.conb.2013.11.00513. S. Sponberg et al., Science 348, 1245 (2015). https://doi.org/10.1126/science.aaa3042
14. T. C. Irving, in Nature’s Versatile Engine: Insect Flight Muscle Inside and Out, J. O. Vigoreaux, ed., Springer (2006), p. 197;
N. T. George et al., Science 340, 1217 (2013). https://doi.org/10.1126/science.122957315. See, for example, C. D. Williams et al., Proc. R. Soc. B 280, 20130697 (2013). https://doi.org/10.1098/rspb.2013.0697
16. P. C. Foster et al., J. Exp. Biol. 217, 2089 (2014); https://doi.org/10.1242/jeb.093021
N. Gravish et al., Phys. Rev. Lett. 115, 188101 (2015). https://doi.org/10.1103/PhysRevLett.115.18810117. J. Aguilar et al., Rep. Prog. Phys. 79, 110001 (2016). https://doi.org/10.1088/0034-4885/79/11/110001
18. G. V. Lauder et al., J. Exp. Biol. 210, 2767 (2007); https://doi.org/10.1242/jeb.000265
M. H. Dickinson, F.-O. Lehmann, S. P. Sane, Science 284, 1954 (1999). https://doi.org/10.1126/science.284.5422.1954
More about the authors
Simon Sponberg (sponberg@gatech.edu ) is an assistant professor of physics and of biological sciences at Georgia Tech in Atlanta.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}