Lubrication of articular cartilage
DOI: 10.1063/PT.3.3898


Imagine supporting a 1-ton weight on your hand and then sliding it along the palm with a slight push of a finger. Pulling off that trick would require, roughly, the level of lubrication that is provided by cartilage surfaces in the major joints of our bodies. Those joints, which enable rotation (or more precisely, articulation) at shoulders, elbows, hips, and knees, are remarkable structures. Indeed, the articular cartilage layers that coat the ends of the bones and slide past each other as we flex our joints are the most efficiently lubricated surfaces in nature. No manmade material can match the ultralow sliding friction, which is a consequence of the lubrication, that cartilage provides at the high pressures and low velocities that our joints experience. Such low friction is essential for their health, as they withstand varied harsh and complex loading, day after day, over a human lifetime.
What is the origin of the lubrication that exists between cartilage surfaces? And can we exploit our knowledge of it to alleviate common and debilitating joint diseases? One example is osteoarthritis, a condition in which high friction is associated with cartilage degradation. Both questions are topics of current research, not least because osteoarthritis imposes a huge burden on society, 1 affecting some 30 million people in the US alone. Deeper insight into how cartilage lubrication relates to disease progression may lead to treatments that ameliorate the condition.
The layers coating our joints
To appreciate the frictional processes that occur at a cartilage surface, consider the basic anatomical structure and composition of our major joints, also called synovial joints. As shown in figure
Figure 1.

The structure of cartilage. A synovial joint (a) is composed of opposing bones that slide and rotate with respect to each other. To support and lubricate those motions, the bones’ ends are covered with a few-millimeters-thick layer of smooth articular cartilage (blue) embedded in synovial fluid; the fluid fills a joint cavity enclosed by the synovial membrane. (b) The cartilage is organized into four main zones. A few tens of microns thick, the superficial zone exposes the articulating, lubricated surface. Water makes up about 70% of cartilage tissue by volume, and cells known as chondrocytes, which produce all the molecules that make up the tissue, occupy less than about 5% of it. (c) The outer boundary of articular cartilage is composed of numerous macromolecules. Among ones that are thought to participate in lubrication are hyaluronic acid (HA), lubricin, and aggrecans. Lipids, much smaller than those macromolecules, are ubiquitous but not shown. (Adapted from ref.

Cartilage possesses four distinct zones, shown in figure
The main macromolecules that permeate the collagen network and have been implicated in its lubrication include proteoglycans, polymers, noncollagenous proteins, and lipids.
2
Proteoglycans are highly charged, bottlebrush-like molecules. In cartilage they are called aggrecans. The ends of those molecules are attached to hyaluronan—also known as hyaluronic acid or HA—a long, linear, negatively charged polysaccharide. The composite structure is shown schematically in figure
Another molecule implicated in cartilage lubrication is lubricin, a flexible, rodlike protein with a charged, carbohydrate-rich backbone. 3–5 Together, phospholipids, HA, and lubricin reside both in the synovial fluid and in the superficial zone of the cartilage. Importantly, phospholipids, in which two hydrophobic chains of CH2 monomers are attached to a hydrophilic phosphocholine head group, form the largest group of lipids in cartilage and its surrounding fluid, the so-called PC, or phosphatidylcholine, lipids. 6 As discussed below, such lipids have been proposed as central to the low friction experienced by flexing joints.
Articular cartilage contains a low density of specialized cells, called chondrocytes, which synthesize the components of the cartilage itself. As the only cells in the cartilage, they are central to cartilage and joint well-being. Crucially, chondrocytes are sensitive to mechanical stresses through a process known as mechanotransduction, which affects how the cells’ genes respond to the stresses. Chondrocytes thus require pressure—or more accurately, normal stress—to function optimally and produce cartilage-maintaining molecules. 7 When subjected to shear stress, however, the cells respond very differently: They overproduce enzymes that may degrade the cartilage network or the molecular components permeating it. 8
That undesirable effect makes the chondrocytes especially vulnerable to high friction. It also provides a direct link between the health of the cartilage and the level of lubrication at its surface. The low friction of healthy cartilage ensures that there is little shear stress at its surface during articulation. However, if the friction increases unduly—due to a traumatic event, such as a sporting accident, or with age—the higher shear stress at the cartilage surface leads to higher shear strain on the chondrocytes embedded in it. That strain in turn leads in a self-reinforcing manner to overall wear of the tissue and eventually osteoarthritis, as illustrated in figure
Figure 2.

The transition from healthy to diseased cartilage. The shear strain on cartilage cells—chondrocytes—is linearly proportional to the product of the friction coefficient μ and the applied pressure. (a) An increase in the friction at the boundary between two surfaces sliding in opposite directions (arrows) can degrade the cartilage through the increase in shear experienced by embedded chondrocytes. The cells respond to increased strain in each panel (from left to right) by producing more cartilage-degrading enzymes (tick marks), which further increases the friction and can lead to osteoarthritis. (Adapted from J. Klein, Nat. Rev. Rheumatol., in press.) (b) A thin histological section normal to an articular cartilage surface shows healthy tissue. (c) An osteoarthritic section exhibits fissures that extend from the cartilage surface into deeper zones. (Adapted from K. P. H. Pritzker et al., Osteoarthritis Cartilage 14, 13, 2006, doi:10.1016/j.joca.2005.07.014 .) (d) This schematic of a knee joint shows degraded cartilage in fully developed osteoarthritis. (Courtesy of stoneresearch.org .)

Frictional energy dissipation
Friction arises from energy dissipation as two surfaces slide past each other, and lubrication is the process by which that dissipation is minimized. The main dissipation processes on cartilage surfaces concern fluid-film and boundary frictions. Fluid-film friction is the viscous dissipation when two surfaces separated by a thin liquid film slide past each other. For a sliding velocity v across a roughly uniform film of thickness D and effective viscosity η, the surfaces experience a shear stress per unit area σ ≈ (v/D)η. In that case, the friction depends linearly on the sliding velocity.
Boundary friction, by contrast, arises when the boundary layers coating the two surfaces are in actual molecular contact. Energy in the form of heat is then dissipated as molecular bonds across the interface between the contacting layers repeatedly form and break as the layers slide past each other. Unlike with fluid films, boundary friction is very weakly dependent on velocity. The upshot: To determine whether sliding is dominated by fluid-film or boundary lubrication, one examines how its friction depends on sliding velocity.
That boundary friction takes place at the slip plane between surfaces has another consequence: The friction is largely independent of the underlying substrates, which are not themselves in contact. Rather, it depends mostly on the structure of the contacting boundary layers. Experiments that measure friction between model substrates coated with given boundary layers can thus provide insight into the friction between living cartilage, so long as the model substrates are coated with similar boundary layers.
Friction in joints
Frictional processes between boundary layers are complicated, and understanding them requires knowing the detailed molecular interactions. Even so, one can use the sliding friction coefficient μ as a descriptive surrogate to quantify their effects. The ratio of the force required to slide two surfaces past each other to the load compressing them, μ is a simple scalar and contains little direct information on the complex physics of energy dissipation. It is, however, useful as a comparative measure between different systems and experiments.
Measurements in synovial joints reveal friction coefficients 2 between roughly 0.001 and 0.03. The weak velocity dependence of friction in such measurements suggests that the dominant mechanism is boundary lubrication, with outer boundaries of opposing cartilage layers in molecular contact. Moreover, by placing sensors in artificial hip implants in living subjects 9 or inserting pressure-sensitive paper between cartilage surfaces in cadaver joints, researchers have measured local pressures as high as 10–20 MPa between cartilage surfaces. The hypothetical scenario that opens this article—a 1-ton load distributed on the palm of one’s hand—would produce a pressure of roughly 1 MPa and is thus a realistic analogy of what our joints experience; and for a surface with a μ of 0.001, a 10-newton nudge from a finger is enough to slide the huge weight.
When the friction between cartilage surfaces is so low, measuring its value in living joints is extremely challenging. Apart from the practical difficulty of manipulating the limbs of living animals to order, the distortion of tissues and ligaments associated with flexing a joint also dissipates energy. Those viscoelastic energy losses manifest themselves as a resistance to the joint’s motion and can mask the very low frictional dissipation from cartilage–cartilage sliding.
Most measurements have been carried out on cartilage in whole joints or excised from joints—so-called explants. Alternatively, researchers have used model surfaces whose boundary layers mimic those on living cartilage. 2 , 5 Both methods have advantages and disadvantages. Cartilage explants more closely reproduce the mechanical properties and surface topography of articular cartilage, but the precise molecular structure of their outer boundary layers is difficult to control and may differ significantly from that in living joints. Model substrates such as glass or mica, on the other hand, have the advantage that any molecular layers attached to them can be carefully controlled. And the controllable attachment of suitable boundary layers can provide molecular insight into boundary friction, even when the substrate differs from cartilage itself, as explained in the last section.
Measuring and modeling
Efforts to understand the lubrication of natural joints date back to the 16th century, when Swiss physician Paracelsus described the oily nature of the synovial fluid in the joint cavity. The complex structure and composition of cartilage, including its high water content, make both fluid-film and boundary processes likely contributors to the overall frictional dissipation. The earliest models were based on lubrication in machines, whereas later ones were based on the fluid films exuded by cartilage between its sliding, pressurized surfaces. In that second class of models, the fluid between the surfaces reduced the friction and supported much of the load via its own pressure. 10 , 11 Most frictional dissipation in joints arises from boundary contact, however, and most research has striven to identify the characteristic structure and molecular composition of the boundary layer at the cartilage surface.
One molecule identified in the early 1930s as a possible lubricating candidate is HA. The charged polysaccharide is largely responsible for the highly viscous nature of synovial fluid and is routinely injected into patients’ joints to relieve symptoms of osteoarthritis. But in 1970, researchers at Harvard Medical School showed that removing the HA from synovial fluid barely affected lubrication, 3 and in later studies, the glycoprotein lubricin was implicated as the lubricating molecule. Additional molecular components of the synovial joint, including PC lipids, aggrecans, and other proteins, have also been proposed as lubricants, either alone or in combination, and as components of the synovial fluid or in boundary layers coating the cartilage. 2
Experimental methods for probing cartilage lubrication have ranged widely—from macroscopic measurement of friction in whole joints or cartilage explant surfaces to microscopic methods. The latter include surface force balance (SFB) methods and colloidal probe scanning techniques, whereby a colloidal particle attached to the cantilever of an atomic force microscope slides past molecular layers on substrates, such as glass. 12 Widely used by our group and others, SFBs can directly measure the normal and frictional surface forces between boundary layers composed of the various molecules of interest, which are attached to atomically smooth mica surfaces. 13 , 14 SFBs provide state-of-the-art sensitivity, force/area resolution, and control over the boundary layers, which can be precisely deposited on the mica surfaces; they also reveal, to within a few angstroms, the thickness of those layers.
Clearly, a prerequisite for understanding the boundary lubrication of cartilage is to reproduce the material’s astonishingly low friction—with coefficients as low as 0.001 at physiological contact pressures of up to 10 MPa. (But keep in mind that a portion of that pressure may be borne by the fluid between the cartilage surfaces.
10
,
11
) Figure
Figure 3.

Plots of frictional force versus normal force as measured by a surface force balance when two mica surfaces slide past each other. The mica surfaces bear boundary layers of either hyaluronic acid (HA, open stars and triangles) or HA complexed with aggrecans (all other symbols). The two cases are sketched to the right of their plotted data. The different curves demonstrate that neither of the boundary layers provides the ultralow friction level measured on articular cartilage, whose friction coefficient is about 0.001 at physiological pressures of 1 MPa or more. (Adapted from J. Seror et al., Biomacromolecules 13, 3823, 2012, doi:10.1021/bm301283f .)

Watery shells around charges
The concept of hydration lubrication, a key finding outlined in the
It turns out that the phosphocholine head groups of the most common phospholipids in our joints, the PCs, are extremely hydrated—that is, densely surrounded by hydration shells. Thus they constitute especially effective lubrication units. Boundary layers exposing close-packed arrays of such head groups—for example, the PC lipid bilayers forming vesicles known as liposomes—have sliding friction coefficients less than 10−4 at contact pressures of 10 MPa or more. 17
Three years ago one of us (Klein) and postdoc Jasmine Seror led a project showing that HA molecules attached to a surface could interact with PC lipids to produce a boundary layer with nearly such low friction,
13
as shown in figure
Figure 4.

Complexes of lipids and a hyaluronic acid (HA) layer exhibit friction coefficients on par with those measured on articular cartilage. (a) In one experiment, PC lipids (blue and green), whose phosphocholine head groups are each joined to two C16 chains, are attached to HA molecules (black). The HA molecules in turn are functionalized with chemical groups (biotin, red) that anchor the HA–lipid complex to a mica surface, which is coated with avidin groups that strongly attract the biotins. (b) In this atomic force micrograph of the complex on the mica substrate, the meandering red lines highlight strings of HA coated with lipids. (c) Experimental data obtained by surface force balance experiments show variations in friction force with load between mica surfaces that are coated with HA–lipid complexes and slid under water (black symbols) or a salt solution (red symbols); friction coefficients μ and local pressures P label select data, and the dashed line shows the mean of the data taken under water. (Adapted from ref.
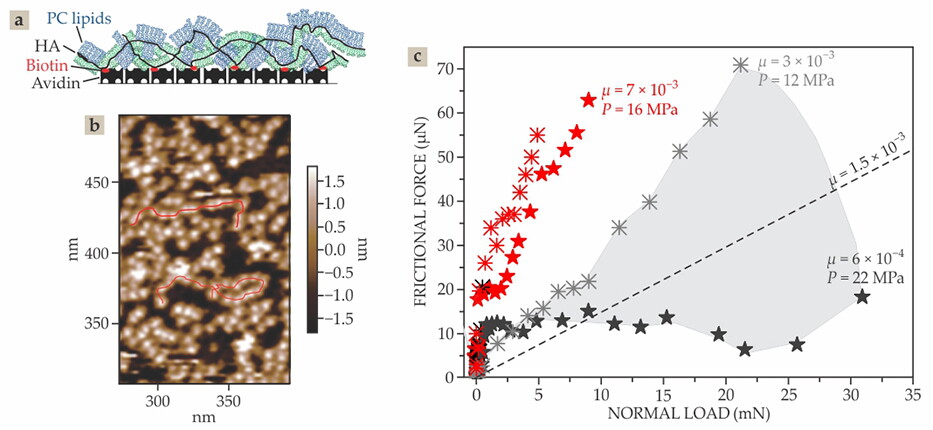
Figure
Figure 5.

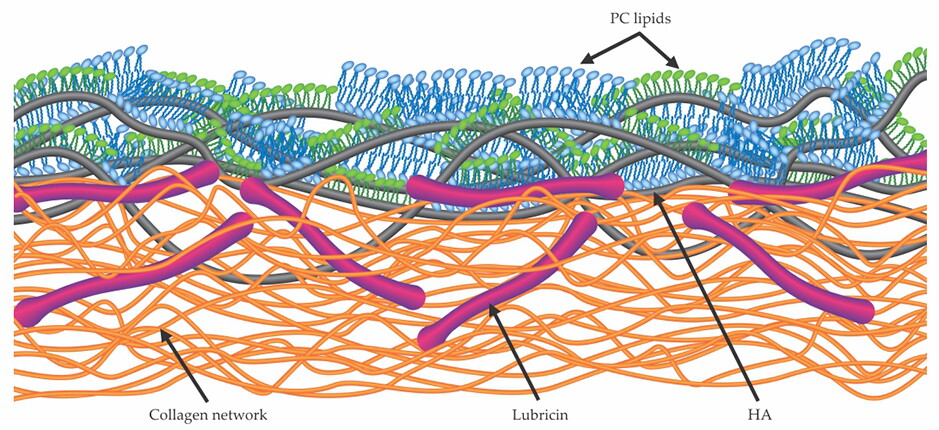
Structure of a proposed boundary layer at the articular cartilage surface. Lubricin molecules (red), known to reside in the outer superficial zone of cartilage and to be attached to its surface, interact with and immobilize hyaluronic acid molecules (HA, black) at the surface. Together, the HA and lubricin molecules form a complex with PC lipids whose outer exposed and highly hydrated phosphocholine head groups reduce friction via the hydration lubrication mechanism outlined in the box on page 52. (Adapted from ref.
Box. Hydration lubrication
Hydration lubrication is an emergent paradigm for understanding friction and lubrication processes in aqueous media, including at articular cartilage surfaces. Because of their large electric dipoles, water molecules in the vicinity of a charge or polar group can surround it to form a so-called hydration shell, as shown in the left panel of the figure (adapted from J. Klein, Nat. Rev. Rheumatol., in press). A large energy is required to break up the shells. And yet individual water molecules in each shell can exchange positions with surrounding bulk water or with water in adjacent shells. The time scale for that exchange can be as short as a nanosecond, depending on the central ion being hydrated.

That combination of being strongly held—and therefore difficult to squeeze out even under a heavy load—while remaining fluid under shear makes hydration shells extremely effective molecular-scale lubricants. And their formation accounts for the extremely low friction provided by hydrated ions and charged polymers, surfactants, and lipids on cartilage surfaces. The hydrated head groups of one class of phospholipid—comprising a negative charge,
Wear between sliding surfaces is inevitable, no matter how well they are lubricated. Thus it is essential that the cartilage boundary layers regenerate themselves. The regeneration is enabled by the fact that the layers’ main components are pervasive in the outer cartilage region and in the synovial fluid bathing the joints. What’s more, because the molecules’ interactions are physical rather than chemical and likely due to charge–charge, charge–dipole, or van der Waals forces, their assembly is essentially spontaneous. The emerging picture of cartilage surfaces—as coated by molecular assemblies that expose continually regenerating layers of robust yet fluid hydration shells—suggests possible new treatments for alleviating osteoarthritis. To name one example, physicians could inject suitable PC lipid vesicles directly into the joint cavity (see figure

JAVIER ROSANO © 123RF.COM

We thank Tonia Vincent, Charles McCutchen, Alan Grodzinsky, and Duncan Dowson for useful discussions and the European Research Council, the Israel Science Foundation, and the McCutchen Foundation for support.
References
1. D. J. Hunter, D. Schofield, E. Callander, Nat. Rev. Rheumatol. 10, 437 (2014). https://doi.org/10.1038/nrrheum.2014.44
2. S. Jahn, J. Seror, J. Klein, Annu. Rev. Biomed. Eng. 18, 235 (2016). https://doi.org/10.1146/annurev-bioeng-081514-123305
3. E. L. Radin, D. A. Swann, P. A. Weisser, Nature 228, 377 (1970). https://doi.org/10.1038/228377a0
4. A. R. C. Jones et al., J. Orthop. Res. 25, 283 (2007). https://doi.org/10.1002/jor.20325
5. T. A. Schmidt et al., Arthritis Rheum. 56, 882 (2007). https://doi.org/10.1002/art.22446
6. M. K. Kosinska et al., Arthritis Rheum. 65, 2323 (2013). https://doi.org/10.1002/art.38053
7. M. Jin et al., Arch. Biochem. Biophys. 395, 41 (2001). https://doi.org/10.1006/abbi.2001.2543
8. T. L. Vincent, Curr. Opin. Pharmacol. 13, 449 (2013). https://doi.org/10.1016/j.coph.2013.01.010
9. W. A. Hodge et al., Proc. Natl. Acad. Sci. USA 83, 2879 (1986). https://doi.org/10.1073/pnas.83.9.2879
10. C. W. McCutchen, Nature 184, 1284 (1959). https://doi.org/10.1038/1841284a0
11. G. A. Ateshian, J. Biomech. Eng. 119, 81 (1997). https://doi.org/10.1115/1.2796069
12. D. P. Chang et al., Soft Matter 5, 3438 (2009). https://doi.org/10.1039/b907155e
13. J. Seror et al., Nat. Commun. 6, 6497 (2015). https://doi.org/10.1038/ncomms7497
14. S. Das et al., Biomacromolecules 14, 1669 (2013). https://doi.org/10.1021/bm400327a
15. B. A. Hills, B. D. Butler, Ann. Rheum. Dis. 43, 641 (1984). https://doi.org/10.1136/ard.43.4.641
16. L. Ma et al., Nat. Commun. 6, 6060 (2015). https://doi.org/10.1038/ncomms7060
17. R. Goldberg et al., Adv. Mater. 23, 3517 (2011). https://doi.org/10.1002/adma.201101053
More about the authors
Sabrina Jahn is a former postdoctoral fellow in the department of materials and interfaces at the Weizmann Institute of Science in Rehovot, Israel. Jacob Klein is a professor in the department and can be reached by email at jacob.klein@weizmann.ac.il .





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}