The speedy secret of shark skin
DOI: 10.1063/PT.3.4460

Shortfin mako sharks are the cheetahs of the open ocean, with some estimates putting their top speeds at more than 100 km/hr. That swiftness may be the result of more than just powerful muscles and a streamlined shape. All sharks have enamel-coated scales, known as dermal denticles, that resemble tiny, translucent teeth, and on fast swimmers like the mako the scales are small—about 0.2 mm, as shown in figure
Friction and pressure
A swimming animal experiences two dominant sources of drag. The retarding force is largely due to viscosity, which in the reference frame of the shark causes water passing over its body to stick to the surface—a phenomenon in fluid dynamics known as the no-slip condition. That condition is responsible for a boundary layer, a thin region of fluid across which the velocity difference between the free stream and the body of the shark is sustained.
Figure 1.

The mako shark is covered with enamel-coated scales, known as denticles. Red areas designate regions on the skin where the denticles can bristle, or flare outward, to angles as high as 30° on the fins and 50° on the flank. Electron microscope images (inset) taken from the flank of the animal show the denticles (a) lying flat and (b) bristled; background scales are unbristled. (c) Grooves jut upward on each scale and are aligned in the direction of the flow of water over the shark. (Photo is by Lukas Walter, shutterstock.com, and inset images are from Philip Motta at the University of South Florida and Maria Laura Habegger, now at the University of North Florida.)

One source of drag is skin friction from the shear stress of viscous fluid rubbing against the surface. Scientists have known for decades that small riblets—stream-wise grooves on top of the scales—reduce skin-friction drag in a turbulent boundary layer by almost 10%.
The second source, known as pressure drag, has the potential to slow the animal to a much greater extent. It is the net force due to pressure around the entire body. Jutting your hand out the window of a moving car illustrates the effect: With your hand held against oncoming air, you can feel high pressure on your palm and low pressure on the back of your hand, with the net force due to pressure against the direction of motion. Streamlining its body—elongating and smoothing out the sharp, angular features—is the mako’s first line of defense against pressure drag.
Pressure drag increases dramatically when the flow separates from the body—that is, when the high-momentum flow of water follows a path that no longer adheres to the shark’s streamlined shape. A wake, or low-pressure region, forms in the gap created by the separation and is accompanied by a reversal in flow direction along the surface. The earlier the flow separates from the body, the larger the wake and the higher the drag. A golf ball has dimples to ameliorate just that effect: Dimples help to keep the flow attached farther around the back side of the ball; the attachment delays the point of separation and reduces the size of the wake. Indeed, a dimpled golf ball can be hit 30% farther than a smooth one. (See the Quick Study by John Eric Goff, Physics Today, July 2010, page 62 .)
Controlling the flow
Unlike a golf ball, a shark has a preferred flow direction—from nose to tail. Figure
Figure 2.

Bristling in action. Consider a single location on the shark’s flank. (a) As water flows (blue) over the body, from nose to tail, the scales (black) lie flat. (b) But as the turbulent boundary layer experiences a higher-pressure gradient, the flow begins to reverse direction (red), which causes the scales to flare outward. (c) That bristling intermittently stops the flow reversal, as the vortices (green) from the flow’s wake are shielded in the scales’ cavities. As forward flow redevelops, the scales lie back down again. That dynamic motion repeats on the scale of milliseconds to minimize pressure drag on key locations of the shark’s body. (Figure by Amy Lang.)
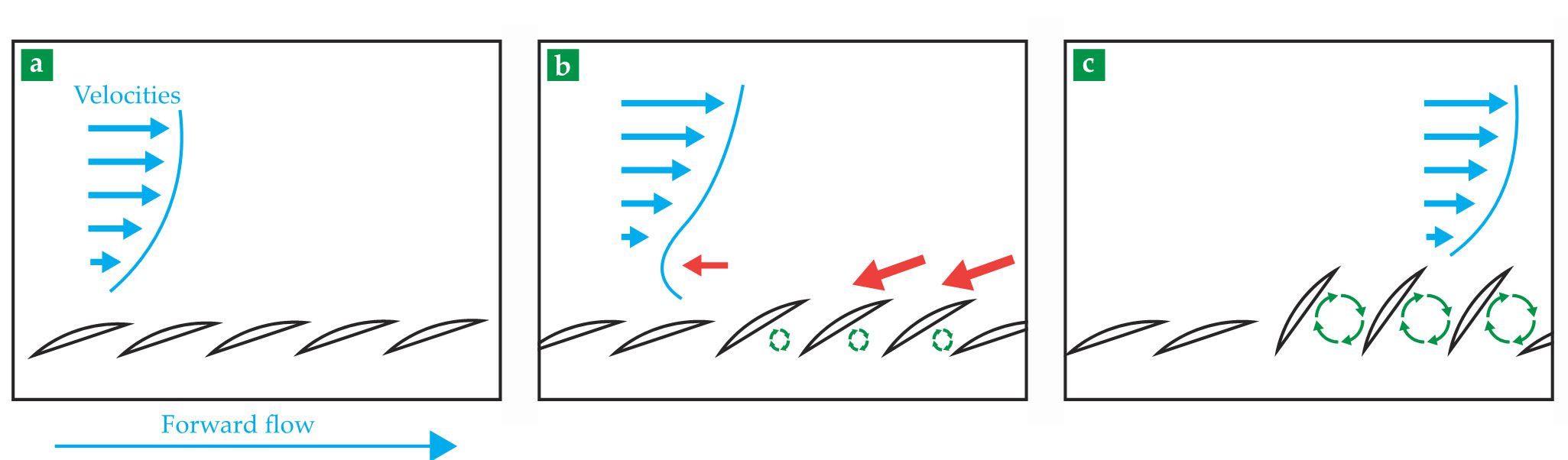
Downstream of that point of minimum pressure along the flank, where the shark undulates in order to swim, the flow experiences an adverse pressure gradient, in that the pressure increases as the flow slows. But because a fluid always moves toward a region of low pressure, the fluid nearest the surface in the boundary layer reverses direction and starts flowing upstream. It is that reversing flow that initiates flow separation. A passive mechanism that inhibits the flow from reversing would prevent, or at least delay, the separation and thus decrease pressure drag.
The scale orientation and bristling capability of the denticles exhibit just that kind of influence. The scales do not bristle as water passes over the skin from nose to tail when the flow is attached. However, on certain body locations—on the flank and trailing edges of the fins and tail, which correspond to regions of high adverse-pressure gradient—the scales sit loosely in the skin and are capable of bristling to angles as high as 50°, but only in the direction of reversed flow, from tail to nose.
In 2012 a collaboration between my group at the University of Alabama and biologists Philip Motta and Laura Habegger (both at the University of South Florida) discovered that the mako’s scale-bristling capability was much larger than anyone had realized. As the flow in the boundary layer begins to reverse (red arrows in figure
A secondary mechanism may also be at work. When scales bristle, they create cavities, whose presence induces turbulent mixing in the boundary layer. That mixing forces higher-momentum fluid in the layer’s upper part (blue velocity vectors in figure
For a description of our experiments to document the ability of mako skin to control flow separation, see the online supplement to this Quick Study.
We hope these findings will inspire the design and manufacture of surfaces that could be applied to submarines and aircraft alike. Flow separation not only produces higher drag; it can also cause airfoil surfaces, such as wings and helicopter blades, to lose lift. Covered with synthetic shark skin, submarines and aircraft would experience reduced fuel consumption, higher speeds, and greater maneuverability.
Online supplement
To document the passive ability of mako shark skin to control flow separation, a collaboration between my group at the University of Alabama and biologists at the University of South Florida took samples from a mako’s flank and mounted them to a smooth, flat plate. The plate was then placed just below a spinning cylinder in a water tunnel to create an adjustable region of flow separation for a turbulent boundary layer forming on the plate. Using a method known as digital particle image velocimetry (DPIV), we measured the water’s velocity field by tracking the motion of seed particles in two sequential images. By time averaging the velocity over 10 000 image pairs, we quantified the percentage of time the flow was reversed—a proxy measurement for the degree of flow separation.
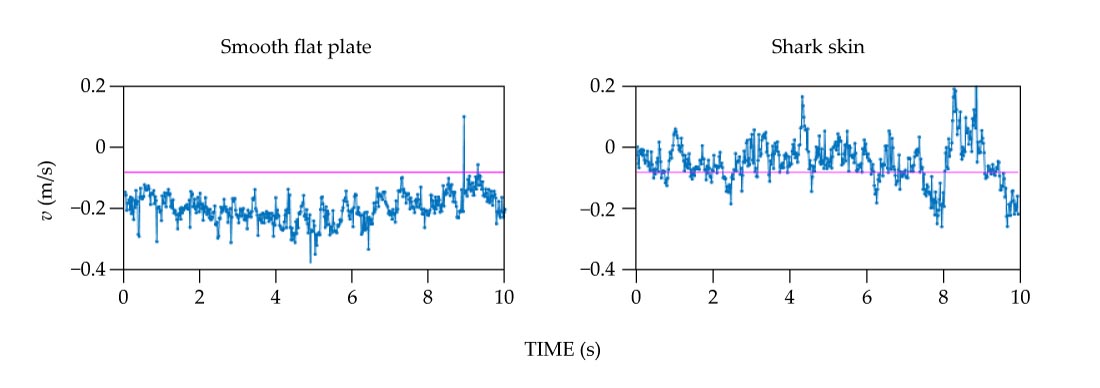
We then repeated the experiment for the same flow conditions but using the flat plate with the shark skin removed. The DPIV data showed that a large region of flow separation that formed over a smooth surface was altered in the presence of the shark skin. Specifically, at one particular point just above the smooth plate, the flow was moving downstream (at positive velocities) less than 1% of the time. By contrast, that percentage increased dramatically, to 73.4%, when measured over the shark skin, as illustrated in figure
Figure S1.

Instantaneous velocity v measured as a function of time. These plots are taken (a) at a point just above the surface of a smooth plate and (b) just above shark skin. The shark skin caused the flow of water over its surface to maintain a forward (positive) velocity 73.4% of the time, whereas the smooth, flat plate experienced reversed flow (negative velocity) nearly 100% of the time. (Data from Amy Lang.)
Although we tested the mako shark skin at much lower speeds than the animal is capable of swimming, subsequent work in a collaborative experiment with biologists has shown that the magnitudes of reversing flow were sufficient to force scales to bristle and inhibit flow separation over the skin.
References
► F. Afroz et al., “Experimental study of laminar and turbulent boundary layer separation control of shark skin,” Bioinspir. Biomim. 12, 016009 (2017). https://doi.org/10.1088/1748-3190/12/1/016009
► D. Bechert et al., “Fluid mechanics of biological surfaces and their technological application,” Naturwissenschaften 87, 157 (2000). https://doi.org/10.1007/s001140050696
► M. Gad-el-Hak, Flow Control: Passive, Active, and Reactive Flow Management, Cambridge U. Press (2006).
► K. Du Clos et al., “Passive bristling of mako shark scales in reversing flows,” J. R. Soc. Interface 15, 20180473 (2018). https://doi.org/10.1098/rsif.2018.0473
► A. Lang et al., “Movable shark scales act as a passive dynamic micro-roughness to control flow separation,” Bioinspir. Biomim. 9, 036017 (2014). https://doi.org/10.1088/1748-3182/9/3/036017
More about the authors
Amy Lang is a professor in the department of aerospace engineering and mechanics at the University of Alabama in Tuscaloosa.





{kind=link}
{kind=link}
{kind=link}