Varying cosmic-ray flux may explain cycles of biodiversity
DOI: 10.1063/1.2800087
Two years ago physicist Richard Muller (University of California, Berkeley) and his student Robert Rohde discovered a 62-million-year periodicity in the wordwide diversity of marine animals. 1 They had undertaken a Fourier analysis of the time variation of the total number of genera listed in John Sepkoski’s authoritative Compendium of Fossil Marine Animal Genera, which begins about 542 Myr ago—when animals first developed the hard exo- and endo-skeletons that facilitate fossilization.
Sepkoski and coworkers at the University of Chicago had been compiling this valuable data set for decades until his death in 1999. Dating the first appearances and final disappearances of tens of thousands of genera depends crucially on age estimates of the stratigraphic layers in which their fossils are found. In 2004 the International Commission on Stratigraphy released a much improved chronology of very old strata based on the best available radioactive potassium–argon dating. “As soon as we applied the new ICS chronology to Sepkoski’s compendium,” recalls Muller, “the 62-million-year periodicity leapt out at us with unambiguous clarity [see figure 1]. It cried out for explanation, but we couldn’t come up with one.”

Figure 1. Fourier spectral analysis of the time variation of the total number of marine-animal genera living at any particular time during the past 542 million years. The prominent peak corresponding to a period of 140 Myr is confidently attributed to the recurring passage of the solar system through spiral arms of the Milky Way.
(Adapted from ref. 1.)
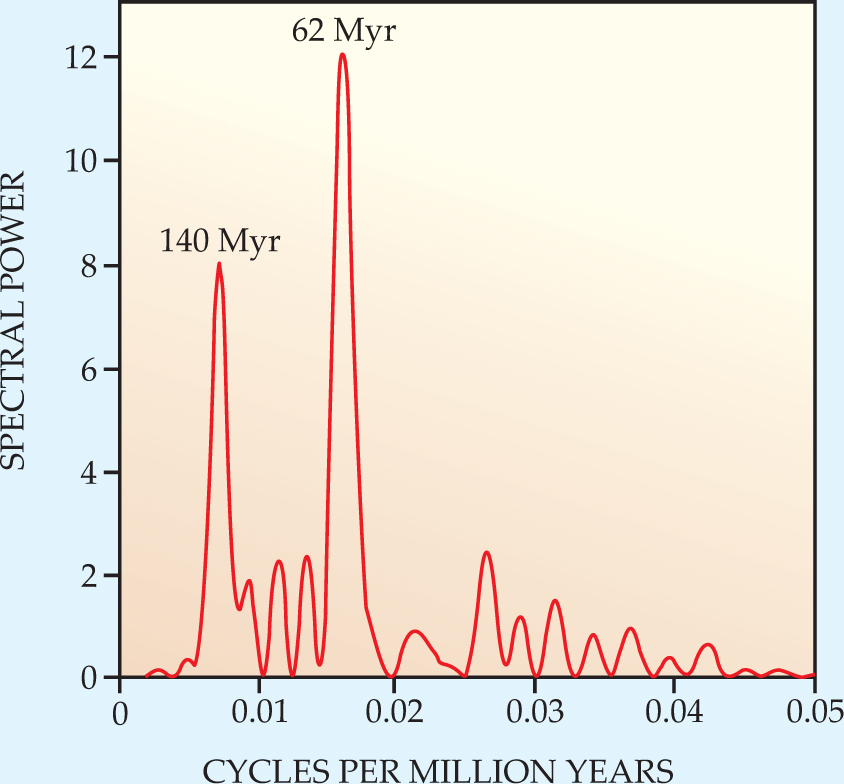
“We had actually been looking for a 26-Myr peak in support of my 20-year-old Nemesis conjecture,” says Muller. Nemesis is the name he gave to a hypothetical faint stellar partner of the Sun that was supposed to pass through the solar system’s Oort cloud once every 26 Myr, loosing a deadly cometary bombardment of Earth (see Physics Today, February 1987, page 17 ).
Extragalactic cosmic rays
Now astrophysicists Mikhail Medvedev and Adrian Melott at the University of Kansas have suggested a cosmic-ray mechanism that might explain the 62-Myr component in the biodiversity variation. 2 It’s long been known that the vertical distance z of the solar system from the midplane of the Milky Way’s disk oscillates with an amplitude of about 230 light-years and a period of something like 62 Myr. That’s the effect Medvedev and Melott invoke as a source of their putatively cyclical cosmic-ray flux variation. The best determination of the mean z-oscillation period 3 from observations plus a model of the galaxy is 63.6 ± 4.4 Myr.
But if the z oscillation of the solar system is so well known, why hadn’t anyone previously suggested it as a source of the biodiversity? Assuming that the neighborhood to the galactic north of the Milky Way’s disk is essentially the same as the neighborhood to the south, any biological effect would have a periodicity of about 31 Myr, a value near which Rohde and Muller found no significant Fourier peak.
But the north–south symmetry of the Milky Way and of the more-or-less spherical halo of gas that envelopes it is in fact broken by the 200-km/s northward motion of our galaxy in the direction of the large Virgo cluster of galaxies. That supersonic motion through the intergalactic medium (IGM) is believed to generate a bow shock front just north of the halo (see figure 2). The halo is formed from the galactic wind of particles that continually blows from the Milky Way. The ram pressure generated by the halo’s motion through the IGM displaces its center to well south of the galactic disk.

Figure 2. The disk-shaped Milky Way galaxy and its gaseous halo, formed from galactic wind, plow northward through the intergalactic medium at about 200 km/s. That movement creates a bow shock front north of the halo, and it pushes the halo’s center well south of the galactic disk. The bow shock and the termination shock front that bounds the halo are places where ambient protons in the intergalactic medium or the halo can, by repeated encounters, be accelerated to become high-energy cosmic rays.
(Adapted from ref. 2.)
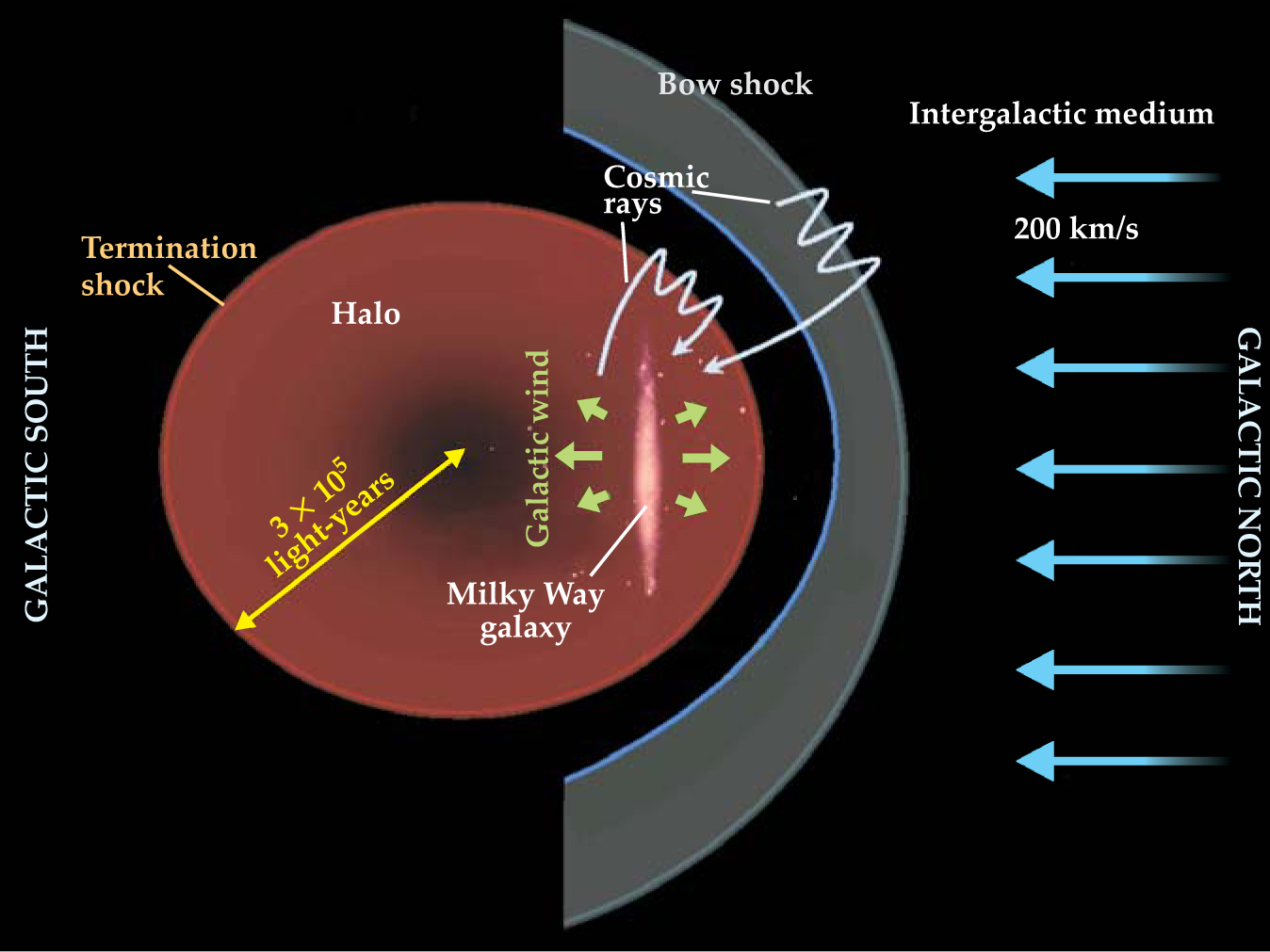
The bow shock and the termination shock front—where the galactic wind finally goes subsonic—are thought to be places where ambient protons and other nuclei in the IGM or the halo are accelerated to become cosmic rays energetic enough (at least 1012 eV) to make their way into the disk against the galactic plasma wind. They are presumed to dominate the flux of cosmic rays reaching Earth from outside the Milky Way itself. Lower-energy cosmic rays that originate inside the Milky Way are generated by essentially the same accelerating mechanism in the much smaller shock fronts of supernova explosions and their remnants (see Physics Today, January 2005, page 19 ).
Given the proximity of the galactic disk’s north side to the termination shock and the bow shock, it comes readily to mind that the solar system might experience a maximum flux of energetic cosmic rays—with possible biological consequences—whenever the solar system is at its farthest northward excursion. At present, we’re only about 30 light-years north of the midplane, but we’re heading north.
How big an effect?
Our motion toward the Virgo cluster and its likely asymmetric effects on galactic shock fronts have been known for some time. But it was not considered a plausible mechanism for the biodiversity variation, essentially because the possible effect on cosmic-ray flux seemed to be too small. Like Earth, the galactic disk is well shielded from cosmic rays by its own magnetic field. The galactic field, extending about 5000 light-years above and below the disk, attenuates the flux of extragalactic cosmic rays by two orders of magnitude. Treating that attenuation with the usual linear diffusion theory of cosmic-ray trajectories in a magnetic field, one would conclude that the flux of extragalactic cosmic rays reaching Earth increases by only about 5% as we go from 230 light-years south of the disk’s midplane to 230 light-years north. It’s hard to imagine that so small a flux variation could have a major effect on biodiversity.
The principal novelty introduced by Medvedev and Melott is their sophisticated nonlinear treatment of the diffusion of extragalactic cosmic rays in the magnetic field that shields the galactic disk. Their Markov-chain computer simulation takes account of the expected large magnetic-field fluctuations, which they presume to be Alfvén waves propagating along field lines. Such fluctuations cause trapping and reflection of cosmic rays with a mean free path of a few light-years.
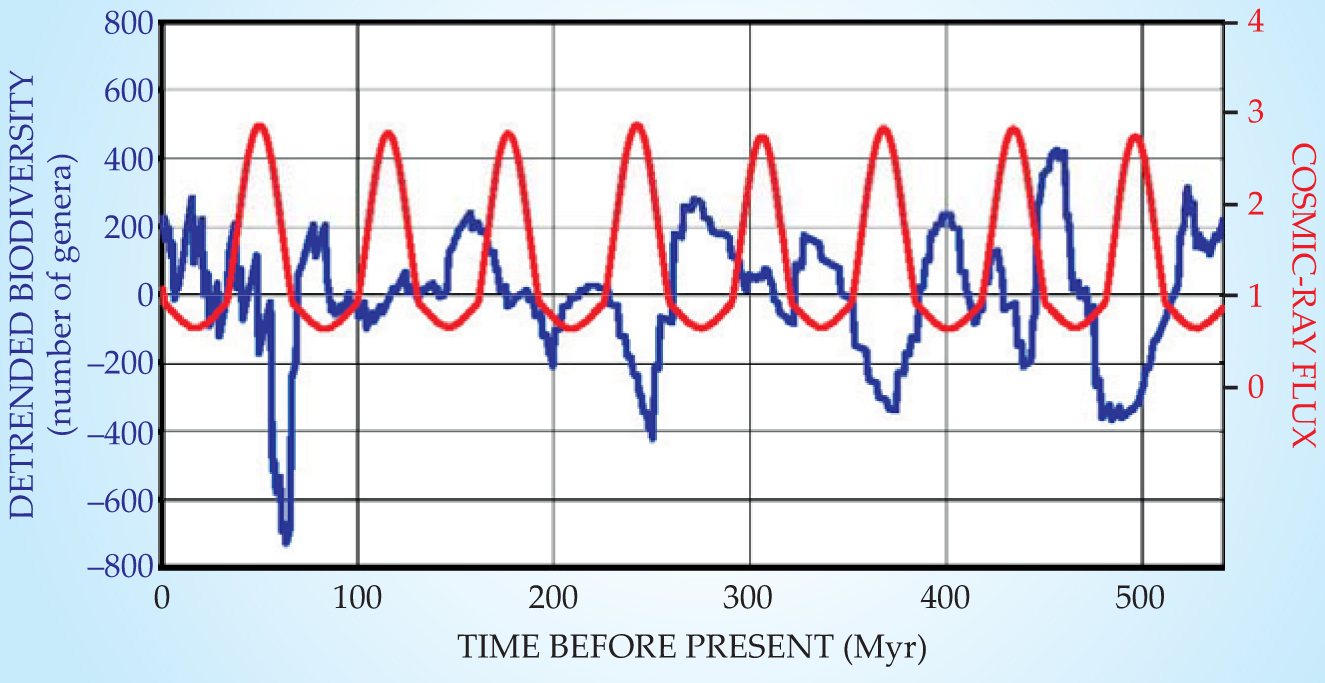
The upshot of the nonlinear diffusion simulation is that the extragalactic cosmic-ray flux increases by almost a factor of five—not just 5%—as the solar system migrates from its southern to its northern extreme. Figure 3 shows the cyclic variation of extragalactic cosmic-ray flux reaching the top of Earth’s atmosphere, as calculated from the Medvedev–Melott model. The extragalactic flux is dominated by protons with energies from 1012 to 1015 eV. Superposed on the calculated flux variation is a detrended Rohde–Muller plot of the number of marine genera over the past 542 million years. Detrending here means subtracting off the long-term smooth variation in the number of genera on time scales much longer than 60 Myr.

Figure 3. The cyclic variation of extragalactic cosmic-ray flux (red) at Earth, calculated from the model in reference 2, is compared with the variation (blue) in the number of extant marine-animal genera.
(Adapted from ref. 2.)
Damaging the biosphere
The model’s nine cosmic-ray flux maxima over 542 Myr correlate suggestively with biodiversity minima. The model has no free parameters that adjust the phase or period of the cosmic-ray cycle to fit the biodiversity data. Medvedev and Melott discuss a number of ways in which a fivefold increase in the flux of extragalactic cosmic rays might kill off lots of marine genera. Cosmic-ray primaries hitting the top of the atmosphere produce showers of ionizing charged particles. What reaches the ground is mostly muons, charged pion-decay products that can penetrate through kilometers of water. The number of muons in a shower increases with the primary’s energy, but the cosmic-ray flux falls off steeply with energy. That tradeoff makes 1012-eV primaries the most prolific source of muons at Earth’s surface. Cosmic-ray muons account for about a third of the average American’s dose of natural ionizing radiation. An increase in the extragalactic cosmic-radiation flux would certainly increase radiation damage to DNA, even among deep-sea dwellers.
The authors also point out that ionization of the atmosphere by cosmic rays can affect climate, electrical storm activity, and atmospheric chemistry: There’s good evidence that ionization facilitates cloud formation, which in turn increases the planet’s albedo. Cosmic-ray ionization near the ground is also known to play an important role in triggering lightning discharges, which produce oxides of nitrogen. The nitrogen oxides create acid rain, and they damage the ozone layer that protects the biosphere from solar UV radiation.
For the moment, Medvedev and Melott don’t advocate any particular one of those mechanisms as the culprit; nor do they argue that the cyclically increased extragalactic cosmic-ray flux is the overwhelming mechanism for biodiversity dips and major extinctions. The great extinction at the end of the Cretaceous Period 65 Myr ago was almost certainly the consequence of an enormous bolide impact. And the even greater Permian extinction 250 Myr ago is attributed primarily to an outbreak of volcanism. Furthermore, local super-novae can produce randomly episodic large increases in the cosmic-ray flux.
“What we do suggest,” says Melott, “is that the increased cosmic-ray flux induces a periodic stress—each time lasting about 10 million years—that increases the vulnerability of the biosphere to whatever else might come along.” The correlation coefficient between the two curves in figure 3 suggests that roughly half the variability of the biodiversity is attributable to the periodicity of the extragalactic cosmic-ray flux. Medvedev and Melott calculate that there’s only one chance in 107 that the observed correlation is a statistical fluke. Of course, the authors do give the correlation a leg up by choosing an astrophysical phenomenon whose period matches that of the biological effect they seek to explain.
“Our hypothesis makes predictions that may soon be put to the test,” says Medvedev. The model predicts that the nearby bow and termination shocks to our galactic north should be prolific distributed sources of 1012-eV cosmic rays, with no comparable source to our south. The most immediately promising observational opportunity, he says, is the scheduled launch early next year of NASA’s Gamma-ray Large Area Space Telescope. Cosmic rays colliding with interstellar gas nuclei produce neutral pions that decay to pairs of gammas. If GLAST looks for GeV gammas coming from hydrogen clouds just outside the disk of the Milky Way, Medvedev and Melott calculate that it should be able to verify or exclude the predicted excess from clouds to the north of the disk.
References
1. R. A. Rohde, R. A. Muller, Nature 434, 208 (2005). https://doi.org/10.1038/nature03339
2. M. V. Medvedev, A. L. Melott, Astrophys. J. 664, 879 (2007). https://doi.org/10.1086/518757
3. D. R. Gies, J. W. Helsel, Astrophys. J. 626, 844 (2005). https://doi.org/10.1086/430250





{kind=link}
{kind=link}
{kind=link}