Single, Physics-Based Model Accounts for the Mechanical Properties of Diverse Diopolymer Gels
DOI: 10.1063/1.2012450
During each heartbeat, the aorta’s walls expand to hold the pulse of blood, then rebound elastically to propel the blood downstream. At rest, a healthy human heart has a systolic blood pressure of about 16 kilopascals. But for strenuous, impulsive effort, like dashing upstairs or hoisting a sack of mulch, the systolic pressure can double, even triple.
If the aorta’s walls responded linearly to such increases, they’d balloon and most likely burst. Fortunately, Nature has endowed the aorta with a nonlinear property called strain stiffening. As stress increases so too does the ratio of stress to strain, the elastic modulus.
The aorta’s strain stiffening arises from its structure, a complex set of concentric, alternating layers of rubbery and stiff tissues. It’s surprising, therefore, to find strain stiffening in the far simpler gels that form the support structures within and between cells (see figure 1).

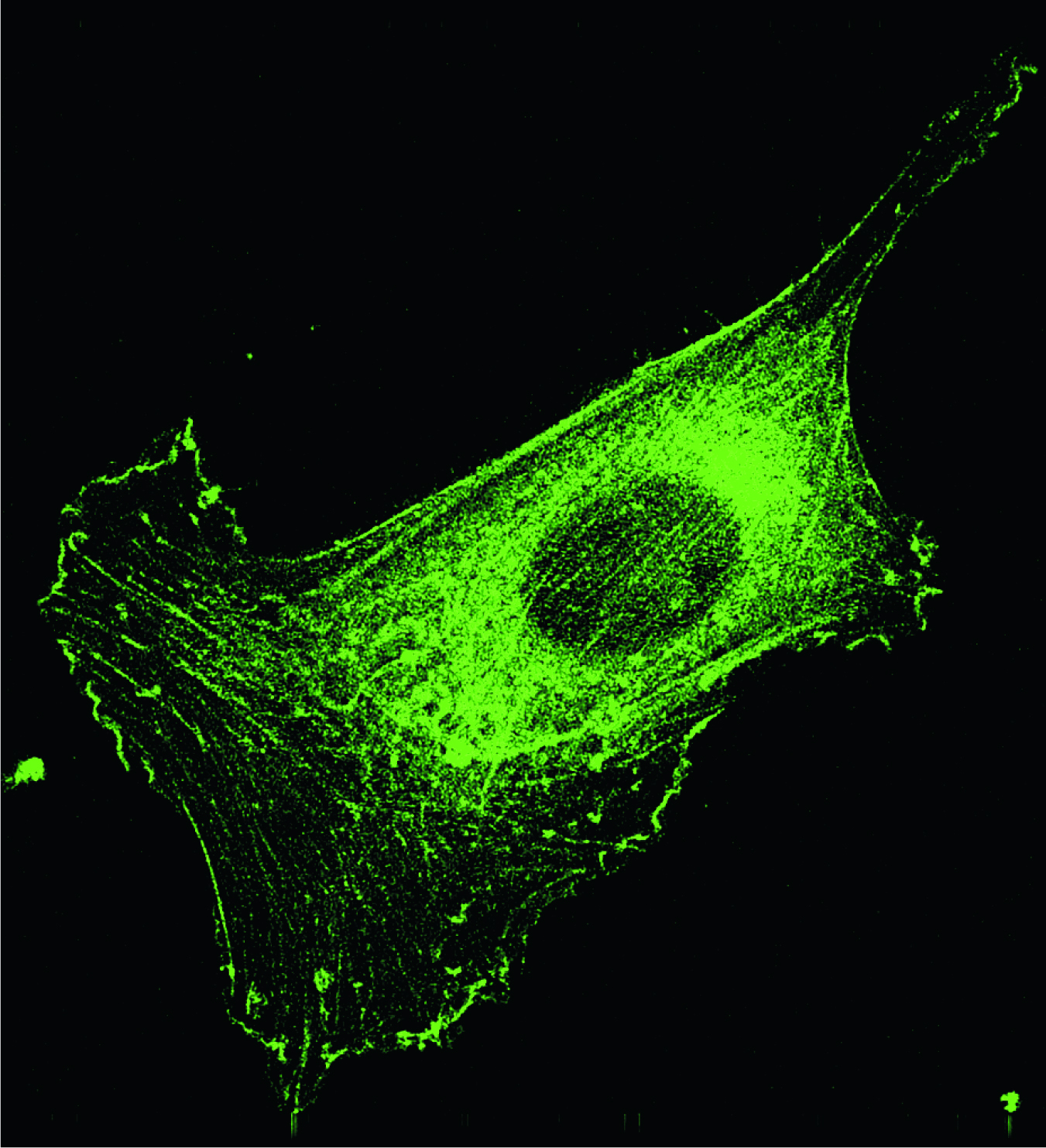
Figure 1. This fibroblast, like other animal cells, is supported by a cytoskeletal network of actin-based filaments, which show up in this fluorescent-tagged microscope image. The cell is about 50 μm long.
(Courtesy of makoto Funaki.)
Those gels consist of weblike filamentary networks of biopolymer embedded in the ambient extracellular or intracellular solution. They perform different functions and bear different stresses and strains. Yet remarkably, despite the variety of composition, strength, and flexibility, the gels’ stress—strain curves follow the same basic shape.
Physicists, when they encounter such similar behavior, look for a universal equation or model. And that’s what Cornelis Storm of the University of Leiden and his collaborators have now found. According to their analysis, a biopolymer gel’s nonlinear response to strain depends principally on the stiffness of its constituent filaments and the density of the links between them. 1 The work suggests that human attempts to mimic certain biomaterials might turn out to be easier than expected.
Storm’s paper builds largely on the earlier work of two of his coauthors, theorist Fred MacKintosh of the Free University of Amsterdam, and experimenter Paul Janmey of the University of Pennsylvania. The other coauthors are also from the University of Pennsylvania: Tom Lubensky, who helped out with the theory, and Jennifer Pastore, who made the measurements the model sought to explain.
Semiflexible polymers
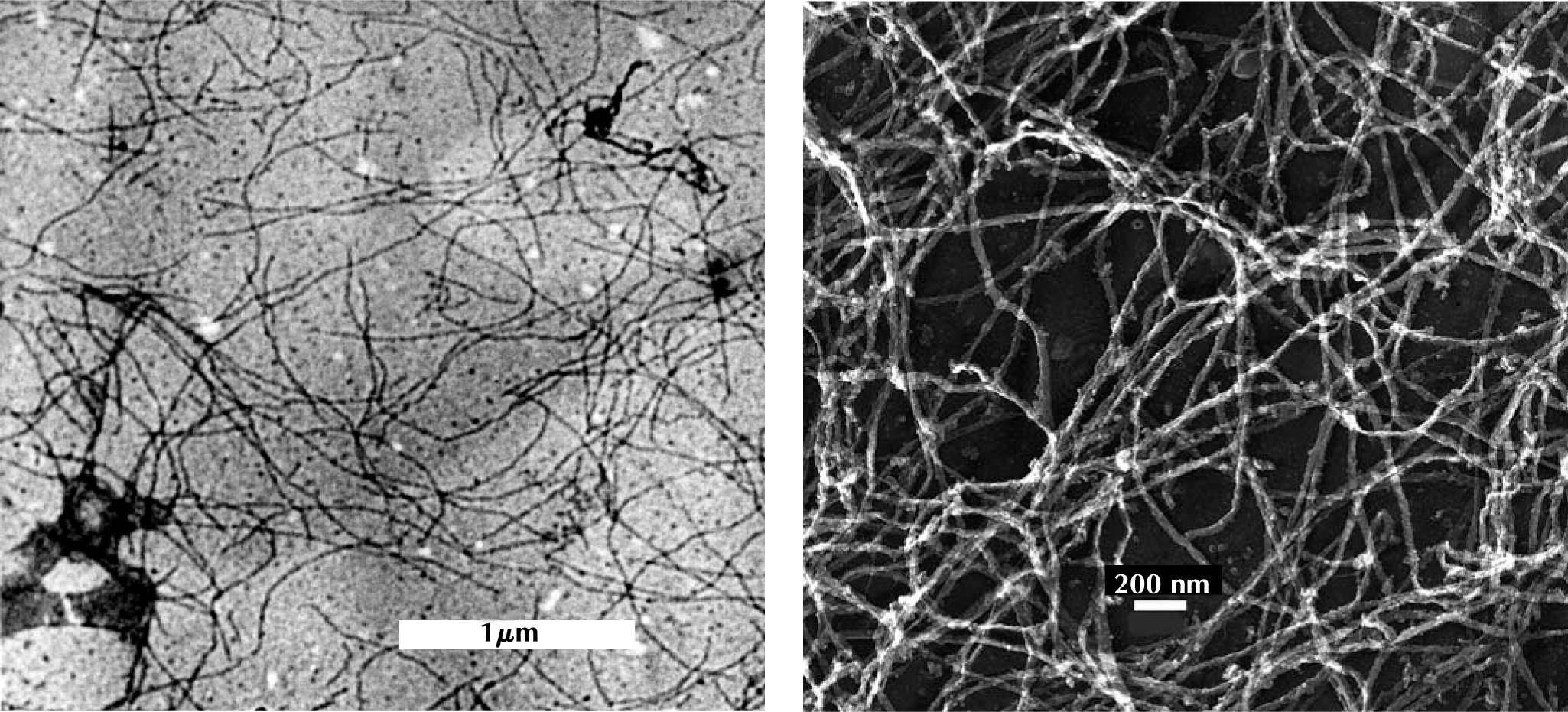
By the 1980s, biologists had identified the main network-forming proteins that animal cells rely on for structural support: collagen and fibrin outside cells and cytoskeletal polymers such as actin inside cells. Close up, as figure 2 shows, the networks resemble spaghetti spread out on a plate.

Figure 2. Biopolymer gels, as seen in these transmission electron microscope images, have similar, open structures. The Neurofilaments on the left are found in nerve cells; the fibrin filaments on the right are found in blood clots.
(Courtesy of Paul Janmey.)
At first, the need for, or even aptness of, a general model for the gels’ mechanical properties wasn’t clear. Actin is much stiffer than collagen or fibrin and could, in principle, be treated as a mesh of semirigid girders. Fibrin, on the other hand, can be stretched to several times its original length before breaking.
Despite the range of stiffness, biopolymers are all what rheologists call semiflexible. That is, the length scales on which filaments undulate thermally and form links with each other are roughly the same. By contrast, the isoprene polymers that constitute rubber wiggle on a scale much shorter than the typical gap between crosslinks.
Semiflexibility was the starting point for a general, physics-based model that MacKintosh, Janmey, and Josef Käs developed ten years ago. 2 Their aim was to account for the large shear modulus and nonlinear response of biopolymer gels.
In the model, the origin of a filament’s elasticity is entropic. Unstrained, a filament can adopt one of many wavy configurations. Stretching it, however, reduces the number of accessible configurations. Like a writhing snake pulled straight by its head and tail, the filament resists extension. The stiffer the filament, or the stronger the snake, the greater the resistance.
To predict the elastic modulus of a network of such filaments, MacKintosh, Janmey, and Käs assumed the network looked, on average, like an open, 3D mesh. That approach could, indeed, account for the low-strain elastic modulus, but, as is often the case, the need for a better model arose when new measurements appeared.
Going nonlinear
Reconstituting biopolymers in vitro is not easy. The protein monomers have to be isolated, purified, and then coaxed into polymerizing without the help of the living cells’ complement of specialized enzymes. Measuring the stress—strain curves of the gels involves placing the samples in a rheometer—essentially two parallel plates, one of which is movable. The rheometer records the stress required to maintain a fixed strain.
In 2001, Janmey and Pastore extended their measurements of seven different biopolymer gels into the nonlinear, strain-stiffening regime to examine the effects of varying filament stiffness and mesh size. At that time, Storm was a postdoc in Janmey’s department. When he saw the data, he wondered how the earlier model could be extended.
He retained the entropic component of the filament stiffness and added a term to account for the elasticity of a fully straightened filament. To model the network, Storm assumed the filaments are distributed isotropically and randomly. Deriving the bulk modulus then boiled down to integrating across the network’s nodes.
That step proved mathematically messy, until Lubensky discovered a fortuitous simplification. If the local deformation matches the bulk deformation—that is, if the deformations are affine, as had been assumed—then certain symmetries lead to a compact, practical expression for the stress tensor.
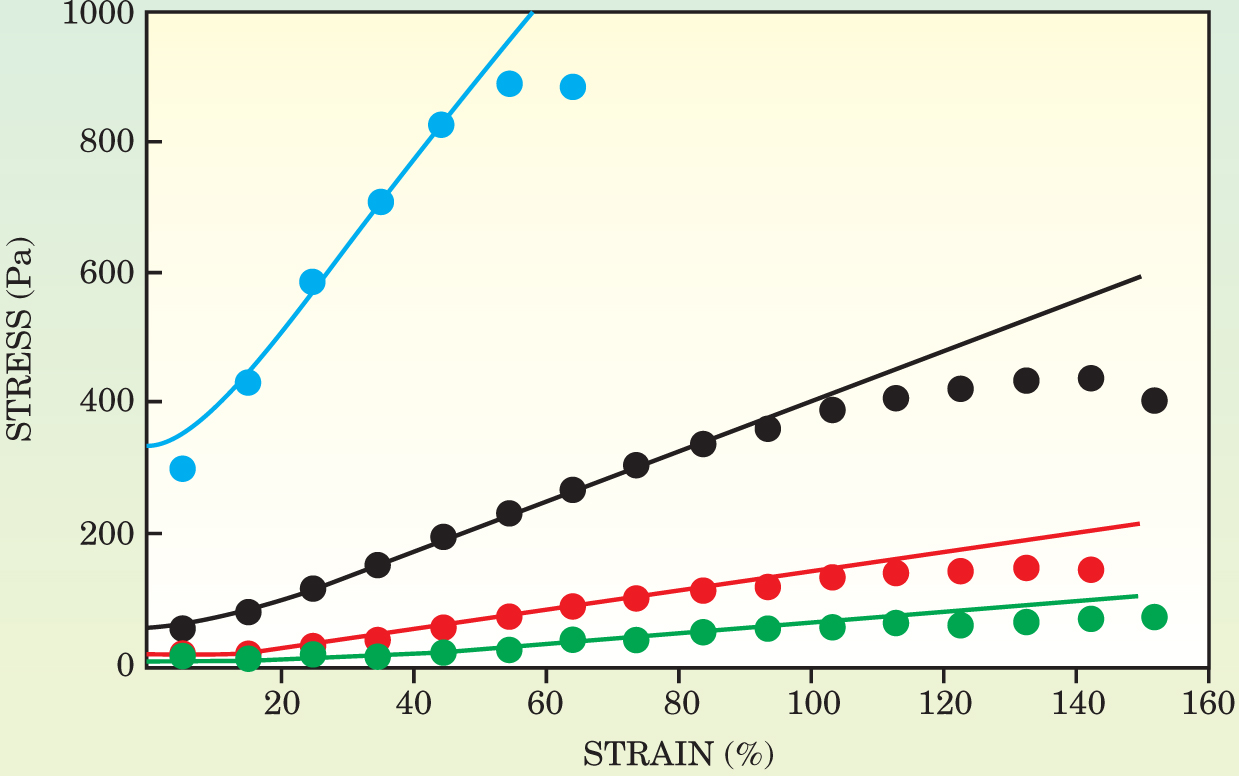
On the scale of a cell, as figure 1 shows, or in low-density regions where inhomogeneities become more important, the affine assumption breaks down. Still, as figure 3 shows, the model proved to account for the data remarkably well. And although the final formula for the bulk modulus has to be solved numerically, it contains just one independent parameter.

Figure 3. The stress–strain behavior of biopolymer gels depends on the concentration of the biopolymer filaments. Here, the data points represent measurements of reconstituted fibrin gels at four different concentrations: 0.5 mg/ml (green), 1.0 mg/ml (red), 2.0 mg/ml (black), and 4.5 mg/ml (blue). As the curves show, the model developed by Cornelis Storm and his coworkers provides a good fit to the data over a range of strains and concentrations.
(Adapted from ref. 1.)
It’s tempting to attribute such apparently universal behavior to a basic requirement identified and picked out by evolution. But, cautions Janmey, that might not be the case for the strain stiffening of biopolymer gels. Some cells, such as long, delicate neurons, do need to withstand high strain, but cells also need to avoid obstructing the free flow of large molecules and even larger organelles. Strain stiffening of biopolymer gels could, perhaps, be an accidental byproduct of their open construction.
References
1. C. Storm et al., Nature 435, 191 (2005) https://doi.org/10.1038/nature03521 .
2. F. C. MacKintosh, J. Käs, P. A. Janmey, Phys. Rev. Lett. 75, 4425 (1995) https://doi.org/10.1103/PhysRevLett.75.4425 .





{kind=link}
{kind=link}
{kind=link}