Microfluidic devices streamline fluorescence experiments
DOI: 10.1063/1.3580484
Louis Sullivan had architecture in mind when he declared that “form ever follows function.” Had his interest been biomolecules instead of buildings, he might have inverted the famous credo—when it comes to biological processes, a molecule’s effect nearly always depends on its shape. An improper protein fold, for example, can spell the difference between a vital organism and a fatally diseased one.
Understanding biological systems, then, rests in part on understanding how biomolecules shift shape. One tool at biologists’ disposal is an effect known as fluorescence (or Förster) resonance energy transfer. In FRET, a donor dye, on being excited by a laser, may transfer energy to an acceptor dye through dipole–dipole interactions. As a result, irradiation of the donor causes both donor and acceptor to fluoresce—each with a distinct color—at relative intensities that depend on the distance between them.
A biomolecule tagged with donor and acceptor dyes thus emits a two-color burst of photons as it drifts through a laser’s focus. The burst, typically gathered via a confocal microscope, hints at the molecule’s conformation: The higher the FRET efficiency—that is, the larger the share of the acceptor’s photons—the closer the dyes. And although early incarnations of FRET relied on ensemble measurements, in 1996 Shimon Weiss led researchers from the Lawrence Berkeley National Laboratory and the University of California, Berkeley, in demonstrating that with an appropriate excitation scheme, FRET efficiencies can be determined for individual molecules.
Sample manipulation in those early single-molecule FRET (smFRET) experiments was often as simple as pipetting a solution onto a slide. With the advent of microfluidics, however, much more sophisticated operations can be realized in the space between a microscope slide and its cover (see the article by George Whitesides and Abraham Stroock, PHYSICS TODAY, June 2001, page 42 ).
Now two research teams have paired that microfluidics technology with smFRET. Ashok Deniz (Scripps Research Institute), Alex Groisman (University of California, San Diego), and colleagues used ultrafast microfluidic mixing to gather high-resolution snapshots of a nonequilibrium protein-folding reaction. 1 Researchers led by Weiss (now at UCLA) and Stephen Quake (Stanford University) automated a high-throughput scheme to investigate effects of chemical environment on RNA transcription. 2 The studies underscore a budding partnership between microfluidics and smFRET.
Into the fold
No one is exactly sure what the protein α-synuclein does. Expressed in high quantities in the human brain, it is thought to facilitate synaptic activity and has been linked with Parkinson’s disease. It is intrinsically disordered in its native state but curls partially into a helix on contact with a cell membrane. Beyond that, the details get murky.
In a 2009 study, Deniz and coworkers at the Scripps Research Institute used smFRET to learn more about the protein’s conformational changes.3 They tagged each protein molecule with an acceptor at one end and a donor near the other; then they measured FRET efficiencies at various concentrations of sodium dodecyl sulfate (SDS), a detergent similar to the phospholipids that make up cell membranes.
The results provided direct evidence for the researchers’ expectations. Absent SDS, FRET efficiencies were moderately high—evidence of a disordered protein in which the donor and acceptor dyes linger, on average, in relative proximity. At finite SDS concentrations, the FRET efficiency either grew or faded, which suggests that donor–acceptor pairs were sometimes drawn closer and sometimes pushed apart as the proteins stiffened into helical structures.
The details of the folding kinetics remained unclear. In principle, insight could be gleaned by mixing SDS into a solution of unfolded proteins and detecting the time-dependent response of the FRET efficiency, but the mixing time would need to be much shorter than the few milliseconds it takes for α-synuclein to fold.
Such ultrafast mixing can be accomplished by squeezing a protein-rich flow between two SDS-rich flows in a microchannel. Provided the flow rates are fast, the protein stream narrows to a length scale small enough to facilitate fast diffusive mixing. Detection of smFRET signals, however, requires slow flow, since each labeled protein must spend enough time passing through a microscope’s focal volume to emit a statistically significant number of photons.
Deniz describes an inherent tradeoff: “Most experiments have either used fast mixing with ensemble measurements or used single-molecule measurements with slower mixing.” Inspired by an idea from then postdoc Yann Gambin, the researchers figured out how to have it both ways.
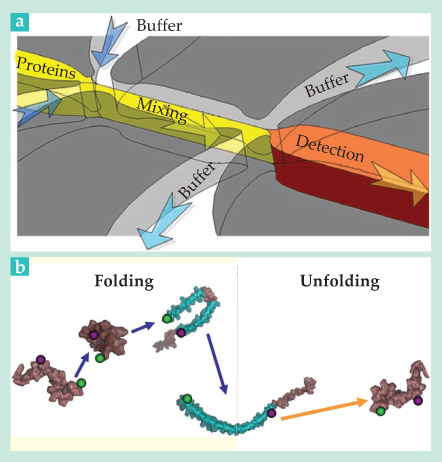
The team designed a device, sketched in figure 1a, in which a protein-rich solution is fed into a mixing channel, flanked by SDS-rich buffer streams. Downstream, after SDS has had time to diffuse into the protein stream, most of the flow is diverted into side channels. Only the protein-rich portion along the centerline passes through a narrow junction into the detection channel, where flow slows dramatically. The quick deceleration allows the first FRET measurements to be made just 200 µs after the mixing event. FRET efficiencies detected at various distances along the detection channel capture the molecules’ time evolution.
The team’s experiments revealed the three-step folding process illustrated in figure 1b. A folding α-synuclein molecule first collapses on itself, then doubles over into a hairpin shape, and only then does it form an extended helix. The entire process is over in about 10 ms. Previous smFRET experiments would have missed it entirely.
FRET on autopilot
Deniz and his colleagues induced α-synuclein folding with a solution of 1.2 millimolar SDS. But what about the other concentrations and conditions that might prevail inside a cell? The question haunts virtually all smFRET studies of biological processes. Rigorous exploration of parameter space often requires a series of titrations and measurements so tedious as to be impractical.
Weiss, for one, suspected there might be a better way—a high-throughput approach like those popular in genomics and drug discovery but amenable to smFRET. Urged by then graduate student Devdoot Majumdar, Weiss proposed the idea to Quake, a microfluidics expert, and the work of devising a high-throughput scheme began.
The collaboration yielded the two-layer microfluidic chip sketched in figure 2a. The bottom layer, shown in red and blue, contains the flow channels: An inlet channel feeds reagents from up to seven different reservoirs into a mixing ring; confocal viewing chambers permit smFRET detection; and two outlet channels dispense with the waste from each run.

Figure 2. High throughput, low maintenance. (a) A microfluidic chip feeds up to seven different reagents into a ring. There, reagents are mixed and Förster resonance energy transfer is detected via a confocal viewing chamber. Automated valves and pumps (black) manipulate the underlying flow with nanoliter precision. (b) Data from a high-throughput experiment show the dependence of RNA transcription on RNA polymerase (RNAP) and glutamate salt concentrations, which vary from 0 to 683 and 0 to 514 picomolar, respectively. (Adapted from ref.
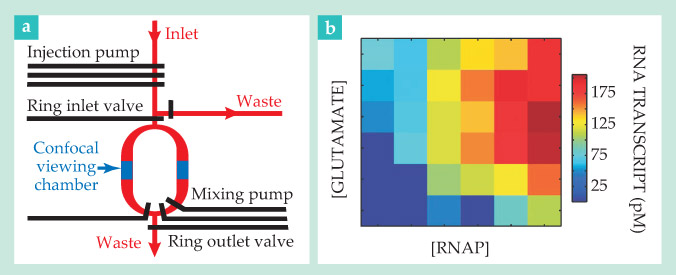
The top layer, black in the figure, houses the controls. Increased pressure in a control channel causes an elastic valve to expand and close off the underlying flow; three such valves in series, closed and opened in succession, constitute a peristaltic pump of nanoliter precision.
The microfluidics aspects of the work weren’t entirely new. In 2009, researchers at Caltech and the Scripps Research Institute used a conceptually similar approach to automate a multistep biosynthesis experiment. 4 Weiss and company’s innovation was to build a device that could accommodate the ultrasensitive smFRET measurements and to write control and detection algorithms that could automate them.
Context is everything
Bacterial RNA transcription is just the kind of process that can’t be roundly characterized with just a measurement or two. It’s reliant not only on RNA polymerase, the enzyme that copies DNA, but also on glutamate, a salt thought to help RNAP molecules escape from their binding locations on DNA strands.
As a proof of concept, Weiss and company used their high-throughput chip to detect transcription rates at 36 points in the RNAP–glutamate parameter space. DNA strands tagged with donor and acceptor dyes at either end indicated the transcription rate. Isolated strands, disordered by nature, would yield moderately high FRET efficiencies. But when transcription picked up, the strands would pair with transcribed RNA to form stiff double helices, and the FRET efficiency would diminish accordingly.
The results, shown in figure 2b, suggest that in most cases, adding glutamate serves to increase the transcription rate. At large concentrations of glutamate and RNAP, however, the team found that additional glutamate only slowed transcription. The result wasn’t a complete surprise—an earlier experiment had suggested as much. 5 More notable was that Weiss and company were able to gather all the data within a matter of hours rather than weeks.
To be sure, the high-throughput technique isn’t yet for every experiment. With a mixing time of about 10 s, it would have been poorly suited to the investigation of nonequilibrium α-synuclein folding. Still, Weiss envisions it as a useful tool for elucidating complex processes such as eukaryotic transcription, which can involve hundreds of proteins.
“Looking at many different conditions used to be tedious and imprecise work,” says Weiss. “Now we just load everything, press enter, and when we come back the next morning we have all the answers.”

Figure 1. Watching a protein fold. (a) A microfluidic chip mixes a protein-rich stream (yellow) with two buffer streams (blue arrows) to initiate a protein-folding event. Downstream, more than 99% of the flow is diverted into side channels. The decelerated flow in the detection channel facilitates single-molecule fluorescence resonance energy transfer measurements. (b) An α-synuclein protein tagged with donor and acceptor dyes (shown, respectively, as purple and green spheres) was revealed to fold from a disordered state to an extended helix via a three-step process (purple arrows). By contrast, a follow-up experiment, run under conditions that lead to unfolding, suggests the helical-to-disordered transition (orange arrow) is direct. (Adapted from ref.
References
1. Y. Gambin, et al. Nat. Methods 8, 239 (2011).
2. S. Kim, et al. Nat. Methods 8, 242 (2011).
3. A. C. M. Ferreon, et al. Proc. Natl. Acad. Sci. USA https://doi.org/PNASA6 106, 5645 (2009).
4. W. K. Ridgeway, et al. Nucleic Acids Res. 37, e142 (2009).
5. J. D. Gralla, Y. -X. Huo, Biochemistry https://doi.org/BICHAW 47, 13189 (2008).





{kind=link}
{kind=link}