Experiments Investigate the Behavior of Pores in Artificial Vesicles
DOI: 10.1063/1.1583525
In a sense, a simple bacterium is like a soap bubble. Both consist of a fluid enveloped by a thin layer of another fluid. But poke a hole in each structure, and their resemblance ends: The soap bubble pops; the bacterium’s single cell reseals. The origin of this difference lies in the competition between surface tension and its lesser-known one-dimensional analog, line tension.
Line tension arises when molecules have to rearrange themselves at the edge of a hole. The rearrangement has an energy cost, which the molecules pay by forcing the edge to shorten. Because a small fraction of molecules bear the cost, line tension is nearly always feeble. In a soap bubble, surface tension prevails. But in a cell membrane, which feels the same pressure inside and out, line tension is the stronger force.
There are reasons to suspect that line tension plays a role in how cell membranes function. Neurons, for example, communicate by exchanging membrane-wrapped packets of neurotransmitter called synaptic vesicles. After making its way across a synapse, a vesicle somehow fuses with the target neuron and delivers its contents. Whatever the fusion mechanism, a pore has to form in the membranes of both the vesicle and the neuron.
But before one can begin to investigate line tension’s possible contribution to cellular fusion and other membrane processes, one has to measure it. And that’s difficult. Line tension is not only weak, but it also acts quickly and on small scales. In water, pores a few microns across close in milliseconds.
Over the past few years, Françoise Brochard-Wyart of the Curie Institute in Paris, France, and her collaborators have developed techniques to observe and understand pore formation in artificial vesicles. In their latest work, the group examined how adding molecules to the pore edge modifies line tension. 1 Their results fit a thermodynamic formula developed more than a century ago by J. Willard Gibbs. As Brochard-Wyart points out, because such formulae are universal, atom-scale models of specific structures must obey them.
Artificial vesicles
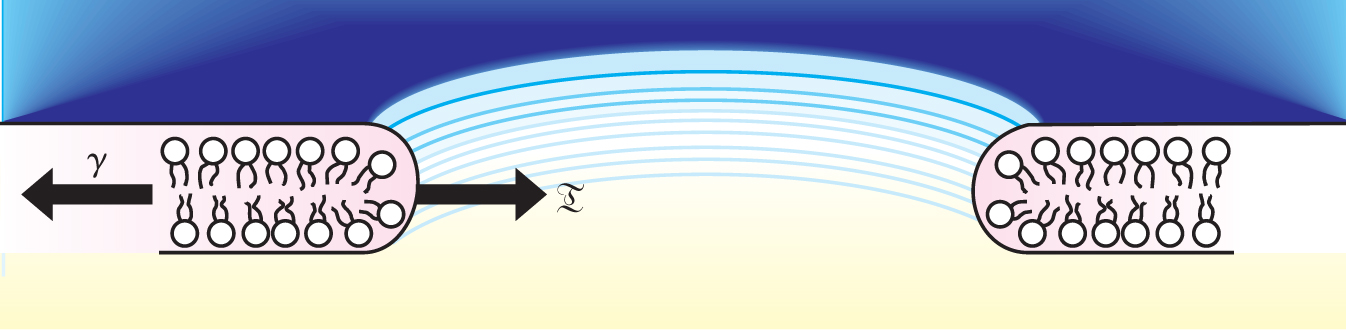
Cell membranes owe their mechanical properties to their main constituent, lipid molecules. Lipids have hydrophilic heads and hydrophobic tails. To keep the tails away from the cell’s aqueous environment and contents, the lipids arrange themselves in two adjacent layers whose surfaces are formed by the hydrophilic heads (see figure 1).

Figure 1. At a pore edge, the hydrophilic heads of lipid molecules are forced further apart than at the rest of the membrane surface. Because this rearrangement costs energy, it gives rise to a line tension
(Adapted from ref. 4.)
Cell membranes also contain specialized proteins and other molecules. But for simplicity, Brochard-Wyart and her collaborators work with vesicles of a pure artificial lipid called DOPC. To make the vesicles, the group uses a technique called electroformation. Dissolved lipids are smeared thickly on two conducting surfaces. After the lipids dry out, the two surfaces are brought to within 1 mm of each other and the intervening gap filled with a water-based mixture.
The two lipid coatings swell and form little blisters that expand across the gap to meet each other. As the blisters merge, lipid molecules break off and spontaneously form vesicles, trapping solution inside. Without intervention, the process takes several days and yields multilayered vesicles with a broad distribution of sizes. But applying an AC electric field of 10 Hz or so across the gap prevents that from happening. As a result—no one knows quite why—double-layered vesicles of the right size for experiments (10–100 µm across) are produced.
A freshly made vesicle flops around in solution like a jellyfish in seawater. Poking a hole in one wouldn’t reveal much about pore dynamics. To tense the vesicles, the experimenters use light. It’s not clear why light creates surface tension, but the result is clearly observable: Under strong illumination, the vesicle membrane swells, stops undulating, and becomes spherical.
Further increasing the illumination stretches the membrane to the point that a pore forms. As the fluid trapped inside the vesicle escapes through the expanding pore, the internal pressure on the membrane drops, as does the countervailing surface tension. When the surface tension approaches zero, line tension asserts itself to close the pore. Figure 1 illustrates the process.
The opening and closing of a pore, if done in water, is too fast to observe on video. Four years ago, Brochard-Wyart and her colleagues realized they could slow the dynamics by increasing the viscosity of the solution that fills and surrounds the vesicles. Adding syrupy glycerol turns a few-millisecond process into a few-second process that can be filmed through a microscope.
By digitizing and analyzing the video data, Brochard-Wyart’s group found that the rate at which the pore radius shrinks is proportional to the radius. Straightforward physics yields a constant of proportionality whose only unknown factor is the line tension. Plotting the log of the pore radius against time, therefore, yields the line tension. For its DOPC vesicles, the group derived 2 a value of 15 piconewtons.
Enter J. Willard Gibbs
One can think of line tension in terms of a quantity called spontaneous curvature. The term applies to amphiphiles: molecules, such as lipids, that have a hydrophilic end and a hydrophobic end. In aqueous solution, molecules of positive spontaneous curvature form curved surfaces that have the hydrophilic ends on the outside. Molecules that have negative spontaneous curvature form surfaces that curve in the opposite sense.
Lipids have almost no spontaneous curvature. Could the line tension be modified by adding molecules of different spontaneous curvature? The Paris group set out to answer that question, which is interesting for at least two reasons: First, adding molecules to weaken line tension could be important for biological processes. In gene therapy, for example, it might ease the delivery of healthy DNA through a cell membrane.
The second reason is that the effects of adding molecules to an edge look reminiscent of the Gibbs adsorption equation. The equation relates γ, the surface tension of the solution, and C, the bulk concentration of the solute, to ΓS, the excess surface concentration. Specifically, the equation states that
where k is Boltzmann’s constant and T is the ambient temperature. The relation between ΓS and γ at a fixed temperature is known as an isotherm.
Brochard-Wyart realized that a 1D version of the formula, in which line tension

To test the 1D Gibbs equation, Brochard-Wyart, Erdem Karatekin, and graduate students Pierre-Henri Puech and Nicolas Borghi measured the line tension of their DOPC vesicles at different concentrations of an industrial detergent called Tween 20. At present, the experimenters can’t vary the temperature in their apparatus, so they measured only one isotherm.
Tween 20 molecules have positive spontaneous curvature. As such, they can be thought of as tiny cones whose hydrophilic heads are larger than their hydrophobic tails. At a membrane edge, the lipids’ hydrophilic heads are forced apart, creating wedge-shaped gaps that the detergent molecules can snugly occupy. By filling the gaps, the detergent molecules alleviate the intermolecular strain and, with it, the line tension.
The results of watching pores open and close at different concentrations of detergent are shown in figure 2. Three distinct regimes appear. In regime I, detergent molecules adsorb preferentially at the exposed edges of the pores. The more molecules adsorbed, the lower the line tension, which drops logarithmically in accordance with the one-dimensional Gibbs formula. The slope of the lower, semi-log plot in figure 2 implies that ΓL is about 2 nm−1.

Figure 2. Line tension at a pore edge decreases in three distinct regimes as the concentration of Tween 20 detergent increases through three critical values: C*, C**, and CMC, the critical micellar concentration. In the top panel, the two axes are linear. In the bottom panel, the concentration axis is logarithmic.
(Adapted from ref. 1.)
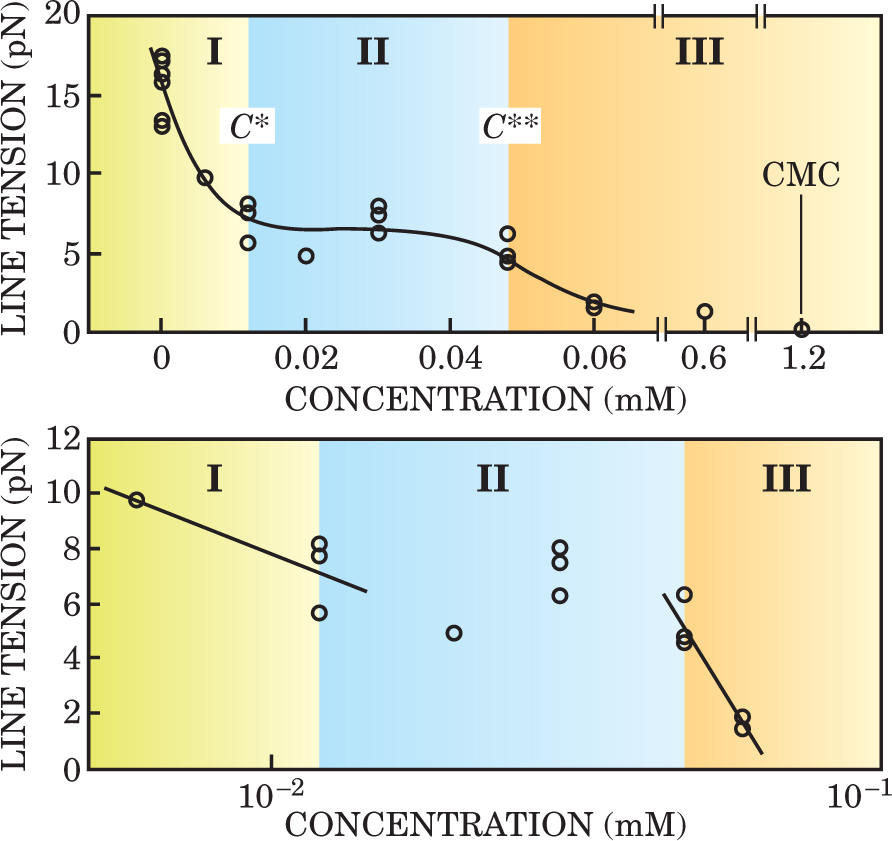
At a critical concentration that Brochard-Wyart labels C*, the line tension stops decreasing. In this second regime, any additional detergent molecules adsorb at the membrane surface, rather than at the edge.
Line tension then stays flat until the detergent concentration reaches a second critical value, C**. At that point, line tension resumes its logarithmic descent, but this time more steeply. In regime III, ΓL is about 9 nm−1. Brochard-Wyart speculates that when C exceeds C**, the detergent molecules resume adsorbing at the edge, but in a new close-packed condensed phase.
Regime III persists until the concentration reaches the so-called critical micellar concentration. At that threshold, the detergent molecules readily aggregate to form spherical structures called micelles. They also dissolve the vesicles.
Having demonstrated that line tension obeys the Gibbs adsorption equation, Brochard-Wyart used the equation to calculate the adsorption energy of a Tween 20 molecule. The derived value, 12 kT, is comparable to the value that Heiko Heerklotz and Joachim Seelig of the University of Basel, Switzerland, obtained three years ago from thermal measurements. 3
In other work, 4 Brochard-Wyart’s group has repeated the experiment, but with cholesterol instead of Tween 20. Cholesterol, whose main biological role is to increase membrane stiffness, has negative spontaneous curvature. As Gibbs might have predicted, adding cholesterol increased line tension.
References
1. P.-H. Puech, N. Borghi, E. Karatekin, F. Brochard-Wyart, Phys. Rev. Lett. 90, 128304 (2003) https://doi.org/10.1103/PhysRevLett.90.128304 .
2. O. Sandre, L. Moreaux, F. Brochard-Wyart, Proc. Natl. Acad. Sci. USA 96, 10591 (1999) https://doi.org/10.1073/pnas.96.19.10591 .
3. H. Heerklotz, J. Seelig, Biochim. Biophys. Acta 1508, 69 (2000) https://doi.org/10.1016/S0304-4157(00)00009-5 .
4. E. Karatekin et al., Biophys. J. 84, 1734 (2003) https://doi.org/10.1016/S0006-3495(03)74981-9 .





{kind=link}
{kind=link}