Teaching Biological Physics
DOI: 10.1063/1.1897564
Over the past few years, people trained in physics and working in physics departments have taken an unprecedented interest in biological problems. A host of new experimental and theoretical techniques has opened up the quantitative study of systems ranging from single molecules to networks of simple agents performing complex collective tasks. Many departments have begun aggressive programs to hire faculty into the emerging field of biological physics. Engineering departments, too, are investing in the interface of the life and physical sciences, both in bioengineering and in related areas such as chemical engineering, solid mechanics, and materials.
Not surprisingly, the new faculty members, like their colleagues, are interested in teaching the subjects that excite them. Meanwhile, physical-science students are beginning to demand courses relevant to the life sciences. And high-level reports such as the National Research Council’s Bio2010 have emerged to stress the importance of quantitative, physics-based thinking for future life scientists. 1 (See also reference and the Reference Frame essay by John J. Hopfield, Physics Today, November 2002, page 10 )
With all this momentum, it may come as a surprise to find that many people, particularly junior faculty, tell us how difficult they have found it to create and sustain new courses in biological physics. We examine some of the reasons for those barriers and offer a few ideas for such courses that we have gleaned from our own and our friends’ experiences.
It matters
The stakes are high. Many physics departments feel that they must struggle to maintain their student enrollments. It often seems that scientifically talented students come to their universities already convinced that their best career options lie in the life sciences. That perception can be frustrating—after all, many exciting advances in molecular and cell biology rest upon physical techniques and ideas. Why don’t students see this and study more physics?
Perhaps it is because when students read course catalogs, they often get no hint of the great ferment going on in our laboratories. Many physics departments teach a collection of undergraduate courses whose outlines are similar to the menu of 30 years ago, despite the fact that the research interests of physics faculty have changed dramatically. The curriculum must change. In particular, we believe that biological physics must become a mainstream course in all physics departments, offered as regularly as, for example, courses on solid-state or high-energy physics. We also believe that many engineering programs would welcome the introduction of such a course, either internally or as offered by physics departments.
Many departments already offer special-topics courses, graduate courses, and seminars relating to biological physics. Unfortunately, such courses often lack the stability enjoyed by a modern physics course, for example. They are frequently electives that busy students can’t fit into their already crowded schedules. Often, the courses have a limited scope, unlike the overviews of a subject that physics departments provide in condensed matter or particle physics. They frequently evaporate when a particular faculty member goes on sabbatical or is needed to teach a more “critical” course. Moreover, they are frequently created from scratch by junior faculty members, who are already heavily burdened.
We believe it is important to offer a consistent, standardized course that is a requirement for at least some flavors of the physics degree, and perhaps for the bioengineering degree. An intermediate-level course can be a good elective for other engineering majors, as well as for majors in biochemistry and even chemistry. We describe ideas for such a course, which can also serve graduate students. As in other intermediate-level courses, students in biological physics have much to learn from a hands-on laboratory, and we describe some of our experiences implementing labs at various levels.
Many of the concepts central to biological physics can be taught at a lower level, and hence in larger service courses. The Bio2010 report urges biology departments to begin requiring rigorous and relevant physics experience. Premedical students are frequently poorly served by traditional physics courses based on the Medical College Admissions Test. We describe some ideas for meeting the challenge of the service course and outline some experiences teaching biological physics in the context of an engineering program.
Now is a good time to look beyond individual courses and consider offering an entire degree program in biological physics, or at least a specialized concentration within the physics major. A recent article discusses ideas along these lines. 3
Six obstacles
Surely many of the assertions we have made are not very controversial. So why do many faculty members find obstacles to creating biological physics courses? We hear professors say that their colleagues offer objections along the following lines.
-
▸ “We don’t have enough people; we’re barely able to teach a complete curriculum of the old stuff.” It’s true. Physics departments have shrunk relative to the halcyon days of yore and have forced faculty members to eliminate many advanced courses they think are important for their students. The only answer is that we physicists must relentlessly prune our offerings to make room for the new, exciting subjects. Do we really need a two-semester sequence in X, when one might do? Do we really need to offer Y every year? Does the enrollment in Z really justify its existence as a regular course, or could it be offered as a reading course? All of those questions amount to: Would we really rather retain X, Y, and Z and miss out on biological physics? They’re not easy questions, but they do admit rational discussion.
-
▸ “We can’t justify a new elective course for our few majors.” This question overlaps the preceding one. We would suggest that the small number of physics majors might result, in part, from a lack of curriculum options that students imagine as relevant to their own future careers. What’s more, we find that biological physics attracts students from several majors, not just a small, captive audience of physics majors.
-
▸ “It’s already offered in another department.” Other departments do offer courses with names like molecular biophysics, physiology, medical imaging, and neuroscience. Closer inspection, however, reveals that the material students need is thinly distributed through several different courses—for example, the several semesters of general chemistry, physical chemistry, organic chemistry, and biochemistry. Students don’t have enough time to take all those courses!
Even more significant is that many courses in other departments turn out to be very different from what we in physics think of as physics education. Our ambition is to give students simple, general-purpose intellectual tools that tie together many kinds of apparently disparate phenomena. Many of those tools are quantitative and involve the modeling of an unfamiliar situation to confirm or falsify some physical hypothesis. We’d like our students to be able to face a problem they have never seen, pull the right quantitative tool out of their bag, and use it to solve the problem. Do you really have a course like that in your medical school?
-
▸ “That’s not really physics.” A course in biological physics can tell a story whose protagonists include Max Planck, Albert Einstein, Marian Smoluchowski, Linus Pauling, Max Delbrück, Hendrik Kramers, Richard Feynman, and a corresponding list of contemporary names. Are they really physicists? Such a course can also serve as an entry to the conceptual foundations of nanotechnology and soft condensed matter. It also can supply a context in which to teach ideas in continuum mechanics, practically banished from many physics curricula (see the Reference Frame essay by Jerry Gollub, Physics Today, December 2003, page 10 ). Is that really physics?
-
▸ “Our students can’t handle it.” We hear this comment surprisingly often. The implication seems to be that hard courses will drive away even the few physics majors left. But students work hard when they think they’re getting something they need. Physical chemistry is a lot of work, it looks good on a medical-school application, and students take it. A course in biological physics can play the same kind of role.
Finally, a worry that comes from within:
-
▸ “I’m nervous! I don’t know all that stuff, and I don’t have time to teach myself and create a whole new course. And what would I cover, anyway? The field is too huge.” We still feel this! It never goes away. But high-energy particle physics is huge, and somehow physicists teach that. In part, we’re comfortable teaching particle physics because we took it when we were students, but more important, it has a fairly stable canon that’s clearly described in textbooks. Standard texts are now starting to emerge in biological physics too, 4 together with the usual ancillary materials like problem sets and solutions. Such curricular materials will make it unnecessary for the overburdened instructor to invent the whole course from scratch.
We have found it surprisingly easy to get a cooperative atmosphere in an interdisciplinary class. The biochemistry majors do catch us making biochemical slips. But if we’re respectful of their own uneasiness with dimensional analysis, then they end up going the extra mile to teach us things we need. Even better, a little hint is often all our students need to team up among themselves in interdisciplinary study groups.
An intermediate-level course
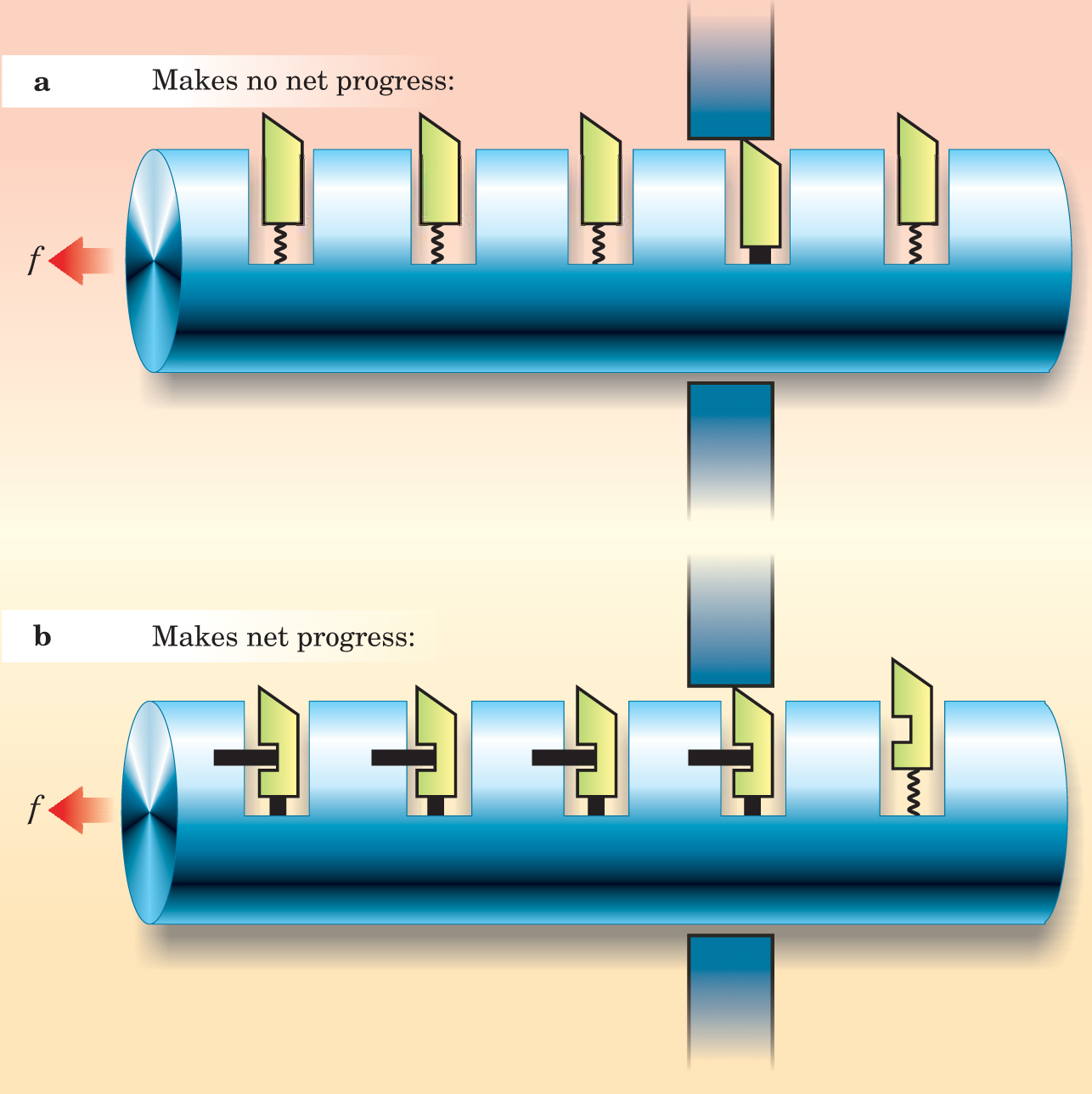
One way to plan a course is to fix the starting and ending points, then find a least-action trajectory connecting them. For example, one of us constructed a course that picks up the story where first-year physics courses typically end and finishes with meaningful discussions of single-molecule manipulation, molecular motors (see figure 1), and the mechanism of nerve impulses, all topics of current research interest. What points must students visit along the way?

Figure 1. Mechanical analogies can illuminate physical ideas sometimes hidden in a mass of molecular details. Naively, it would seem that thermal kicks would drive the ratchet in (a) to the right and do work against a load f. Once the students have worked through the microscopic details of why it doesn’t work, they are ready to study the modified ratchet in (b), where a mechanism releases each pawl only after it emerges on the right side of the wall. The device may seem fanciful, but it contains the essence of an idea currently believed to underlie real molecular motors.
(Adapted from P. Nelson, ref. 4; used with permission of the publisher.)
In a nutshell, the answer was to begin with simple estimates and dimensional analysis, then review a little cell biology to get the players on the stage. Next, students needed a little kinetic theory, leading to the key conceptual foundation of random walks. From that vantage point, one can see many of the phenomena that make the sub-micrometer realm so different from our own—for example, the strange world of low Reynolds number. 5 Students can also get a feeling for the tendency of entropy to increase.
Only when students had a feeling for entropy did the course introduce its abstract definition. But the abstract viewpoint has its virtues, as it leads to a compact and general analysis of entropic forces. Viewing chemical forces as a particular case of entropic forces (see figure 2) was a new experience for most students and one that led naturally to the study of single-molecule devices like enzymes and molecular motors. Students found it satisfying to analyze a case in which an entropic force can be rigorously computed and compared to experiment, as in the mechanical extension of long molecules of DNA. The course culminated with a discussion of the collective behavior that emerges when many similar molecular agents act in concert to create traveling nonlinear waves of excitation—the nerve impulse.

Figure 2. Chemical forces are partly entropic in character. If initially the number of buffalo in the lower-energy state exceeds that state’s equilibrium value, there will be a net flux of buffalo into the higher-energy state. Reasoning in this way gives students a physical intuition for the concentration-dependent part of the chemical potential.
(Cartoon by Larry Gonick from P. Nelson, ref. 44; used with permission.)

The course also assigned reading selections from a mainstream cell-biology textbook. There is no substitute for learning the language, and the iconography, from an industry-standard source, although some beautiful and technically accurate popularizations are also quite useful. 6 Some course assignments also involved the use of computers. Students need to get into the habit of turning to mathematical software to examine the behavior of a model; they also need the habit of turning to a molecular-visualization package and allied online databases to examine a macromolecule of interest.
The syllabus just outlined will not suit everyone, of course. But it does have some claim as a conceptual framework into which many more advanced ideas can be fitted. We think that students need the core ideas of several disciplines before they move on to more abstract topics like neural and metabolic network analysis. Whatever topics you choose to cover, we suggest that every concept be rooted in some quantitative experimental data, the same rigorous standard that is applied when framing a course in, say, quantum mechanics. As physicists, we were taught that if we can write and solve a simple model that draws a curve through some real data and that is rooted in concepts that explained other kinds of experiments, then we may have learned something. Of course, plenty of life scientists concur with such an ethos. We would like to instill it in students, even if it means skipping much of the voluminous factual material of cell biology.
Laboratory courses
Perhaps the simplest way to incorporate experiments in biological physics and soft condensed matter physics into the undergraduate curriculum is to introduce them into standard junior- or senior-level physics lab classes. Consider the current menu of classic experiments—the speed of light, the electron charge to mass ratio e/m, Mössbauer spectroscopy, and so forth. That lineup exists not because each experiment teaches a particular skill that is so important later in a student’s professional life, but because, taken as a whole, it exposes students to the interplay between theory and experiment and incorporates a healthy dose of error analysis. Viewed this way, a lab in which students use an inexpensive laser-tweezer setup 7 to learn about optical trapping and Stokes drag is surely as important as one in which they use a Michelson interferometer to measure the refractive index of gases. Likewise, examining the Brownian motion of micron-size spheres through a microscope and determining Avogadro’s number as Einstein proposed and Jean Perrin carried out must be as important as measuring e/m.
At the graduate level the possibilities are much greater, particularly as departments embark on interdisciplinary education and research efforts such as the Integrative Graduate Education and Research Traineeship (IGERT) program supported by NSF. The experience of one of us with such an effort has provided the resources and students to experiment with how a graduate course can be structured. 8
A centerpiece of that effort is a dedicated laboratory—home to a course taken by all students in the IGERT program—that combines faculty from applied mathematics, physics, neuroscience, physiology, and molecular and cell biology. The students come from the above departments and others such as biomedical engineering and even astronomy. In view of the diverse clientele and a desire to give all students a meaningful laboratory experience, the notion of a methods course that would simply survey current laboratory techniques in biological physics was rejected. Instead, the course emphasizes the interplay of theory and experiment through a case-study format. That serves the students well, for each student comes with definite strengths and weaknesses. Graduate students in applied mathematics, for example, have no problem with the more theoretical underpinnings of the various lab experiments. But they must learn to ask questions with an experiment, a skill that is usually underemphasized in their curriculum. Students in biochemistry are very familiar with biological protocols but are weak on dimensional analysis and the study of the partial differential equations that describe diffusion or fluid flow. Thrown together in a lab course where cooperative learning is emphasized, the students can produce remarkable results.
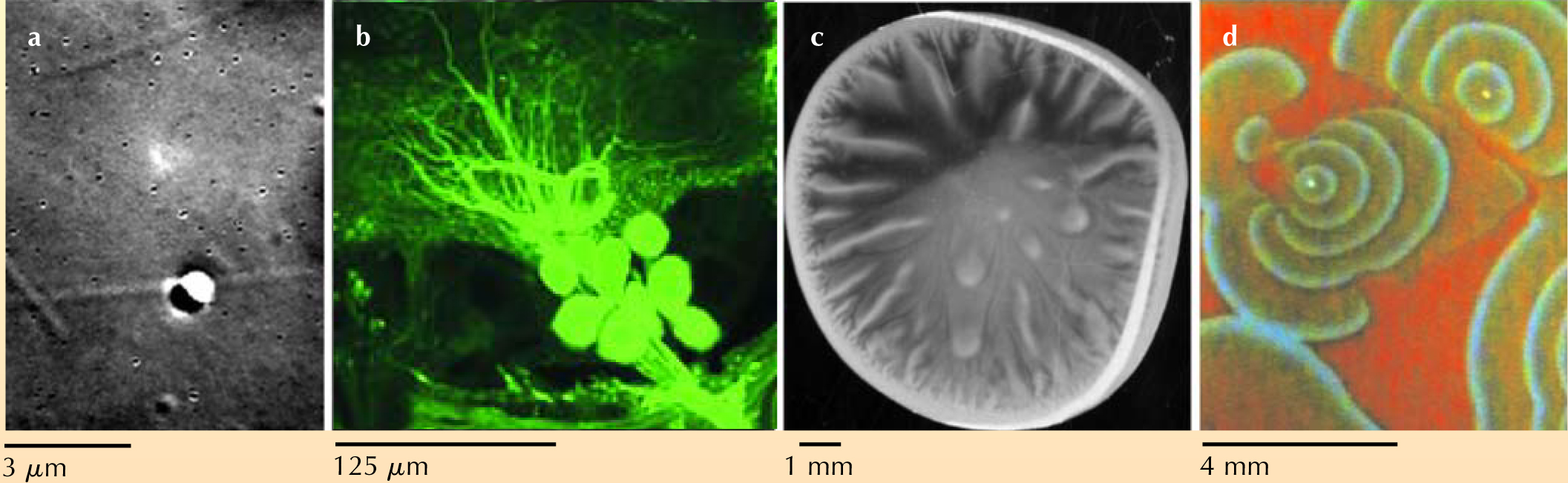
A key issue is determining what are the right experiments. Based in part on the research expertise of the faculty running the course, the decision was made to organize the experiments by length scale, as illustrated in figure 3. After several weeks of introductory lecture material on the essentials of dimensional analysis and estimation, Stokes drag and the Stokes–Einstein relation, electro-physiology, pattern formation, microscopy, micromanipulation, and time-series analysis, each pair of students spent the remainder of the semester on one experiment. For example, at the molecular scale of nanometers, students investigated the properties of the motor protein kinesin as it carries microspheres along microtubules. Related to that study was an investigation of the Kramers problem of thermally assisted hopping over a potential barrier created by two nearby optical traps. At the cellular scale of microns, students measured action potentials in neurons of the moth Manduca sexta. Particle-tracking studies of bacterial chemotaxis and related fluid motions explored scales of hundreds of microns. Experiments at the centimeter scale investigated pattern-forming processes including bacterial bioconvection and the Belousov–Zhabotinsky reaction known for its rotating spiral waves.

Figure 3. Experiments at several length scales are the cornerstone of an interdisciplinary laboratory course for graduate students. The photographs here illustrate (a) a microsphere (large black-and-white structure) pulled along a microtubule by the motor protein kinesin, (b) fluorescently labeled ganglia in the moth Manduca sexta, (c) bioconvection patterns in a drop of bacterial suspension, and (d) spiral waves from a Belousov–Zhabotinsky reaction that takes place in a petri dish.
(Images a, b, and d courtesy of Koen Visscher and Robert Reinking, University of Arizona.)
Each experiment has a well-established underlying theoretical description. Like the experiments themselves, some of those theories are quite involved. Yet, by spending much of a semester focused on both theory and experiment, surrounded by fellow students from a broad range of departments in the atmosphere of a research group, the IGERT students mastered them.
The laboratory layout includes common space for the students to gather for theoretical lectures and to make oral presentations. That space also doubles as the home for an undergraduate biological physics course in which the experimental setups serve as sophisticated demonstrations. What better way to lecture about bacterial chemotaxis than with a suitable microscope setup three meters away so the students can see it with their own eyes?
A no-prerequisites course
A course for students with no university-level physics experience can be structured in many ways. For example, one could add a half-credit seminar that runs parallel to an existing version of first-year physics. One such course was recently taught by Jané Kondev and Robert Meyer at Brandeis University. 9
The course is primarily intended for first-year students who are considering a major in biological physics, although enrollment of premedical students and students from other majors is not uncommon. As a survey, the seminar plays the dual role of attracting students to the major and providing motivation to labor through the daunting mainstream introductory courses in physics, chemistry, and biology. The course meets once a week over two semesters. Thus, the time commitment required of the students is not too big, which is particularly important for freshmen, who are very busy fulfilling other university requirements. The course has no exams, but the students have weekly homework and reading. The homework consists mostly of estimation; the readings are taken from elementary physics and biology texts.
The physics content of the course follows the first-year physics sequence, with mechanics and thermodynamics in the fall semester and electricity and magnetism and optics in the spring. The corresponding coverage has the advantage of letting the students see how physics concepts treated in other courses are applied practically to problems in biology. For example, a simple model of springs connected in parallel leads to an estimate for the stretch modulus of single-stranded DNA. The modulus, in turn, can be read from force–extension graphs obtained in single-molecule DNA stretching experiments.
In addition to applying principles of mechanics and thermodynamics, the course introduces new concepts such as diffusion. The mathematics of diffusion is developed from a one-dimensional random-walk model that also gives insight into the size of polymers in solution and allows for simple estimates of the size of the E. coli genome. For example, students study micrographs that show the bacterium’s DNA spilled out of the cell; those images yield a crude estimate of the radius of gyration. In the spring semester, simple concepts from electricity and magnetism and optics are applied to such topics as conduction by ion channels and the compound lens of a bee’s eye.
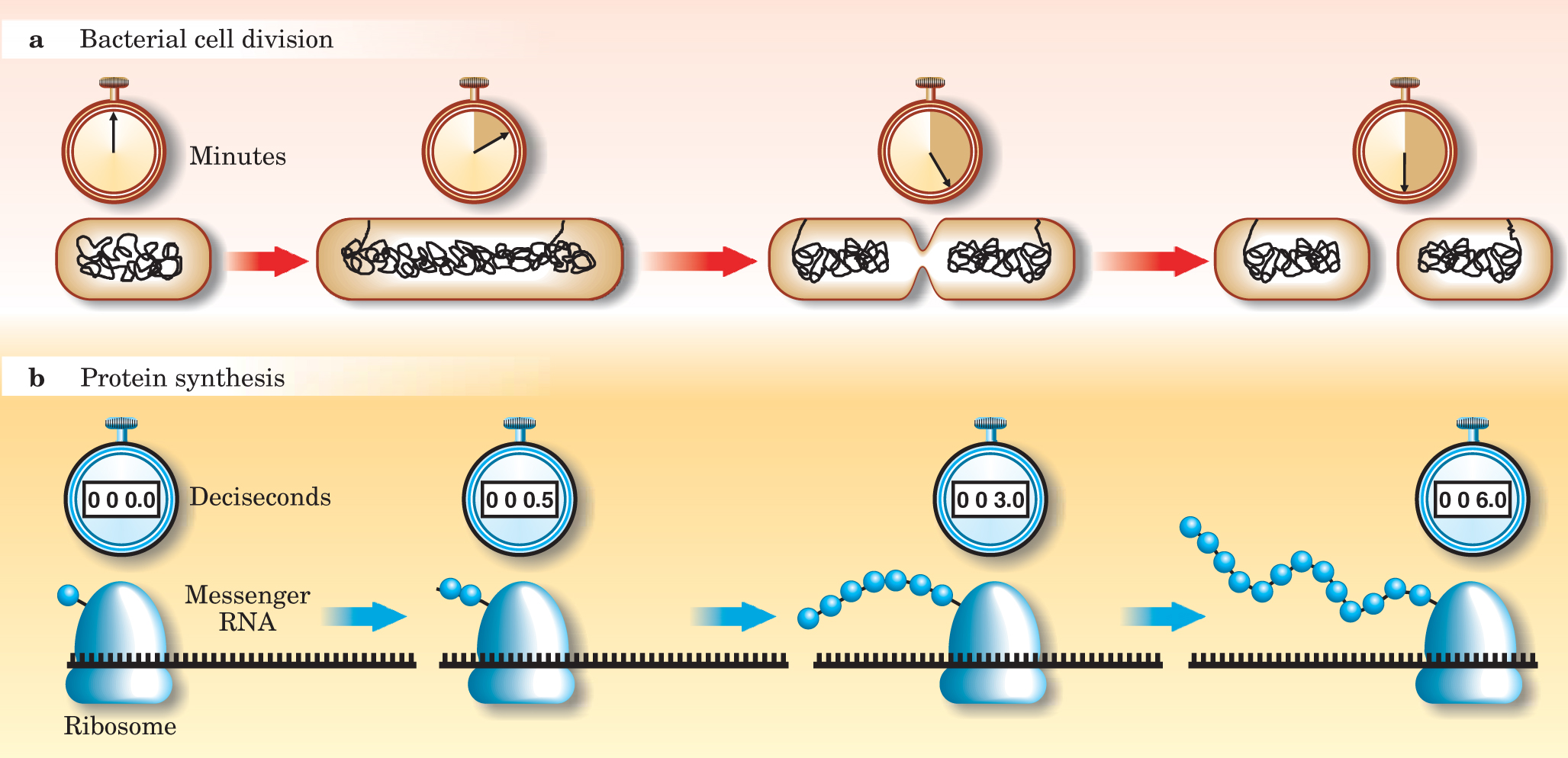
The goal of the course is to introduce the field of biological physics by examining the living world quantitatively. A corresponding course theme is the idea that simple quantitative estimates can reveal all sorts of interesting information. An example is given in figure 4, which shows how the cell division time for E. coli can be used to estimate the rate of amino-acid addition on the ribosome.

Figure 4. The rate of protein synthesis on ribosomes in E. coli can be estimated from the cell division time. As shown in (a), the bacterium divides in about half an hour. During that time, the mass of all the proteins roughly doubles. To estimate the total protein mass, note that about 20% of the cell volume is taken up by proteins, the volume of E. coli is approximately 1 µm3, and the density of proteins is close to that of water. Thus, the protein mass is about 0.2 × 1 µm3 × 1 g/cm3 ⌣ 1011 daltons; a Da is equal to an atomic mass unit. An examination of the 20 amino acids found in living matter reveals that their mass is typically 100 Da. The number of amino acids that need to be strung together to make all the proteins for the two daughter cells is therefore 1011 Da/100 Da = 109. One can estimate from microscopy that an E. coli bacterium has on the order of 2 × 104 ribosomes, so the average rate of amino-acid addition on any one of them is 109/(2 × 104 × 2000 s) ⌣ 20 s−1, as shown in (b), and which roughly corresponds to the experimentally established value.
(Adapted from R. P. Phillips, J. Kondev, Physical Biology of the Cell, to be published by Garland Press. Used with permission.)
Biological physics for engineers
Physics undergraduates and engineering undergraduates typically have different motivations for their courses of study, and those differences must be respected when teaching biological physics to engineers. Broadly speaking, physics students study science to understand nature, whereas engineering students study science because it is the basis for technology. In particular, although engineering students appreciate the importance of fundamental understanding and are capable of the same level of mathematical sophistication as physics students, they are eager to know the application of a model before they are ready to give their full attention to its development. Engineering students want to see the connection of their course-work to the “real world” of industry. But, having studied topics such as beam theory, plate theory, and transport theory in their traditional courses, they are eager to see how that material applies to biology. Bioengineering students are especially excited to see material familiar from their biology courses treated from the physical point of view.
What does the engineering student’s perspective mean for a syllabus? One approach we have found to be successful is to organize the syllabus around biological or biotechnological questions. Engineering students are particularly receptive to questions that relate to optimization or physical constraints. For example, at the beginning of the course, one might discuss the packing of DNA into the head of a virus. A simple question is, How much work does it take to stuff the DNA into the head? A natural place to start is with the random-walk model of a polymer. The students are then led directly to the ideas of entropy and entropic forces. That path has the side benefit of helping to cement the students’ understanding of entropy, which, in the engineering curriculum, is typically first encountered from the abstract perspective of thermodynamics. Once the idea of an entropic force is in place, it is natural to introduce single-molecule experiments and experimental force–extension curves, and then point out the limitations of the random-walk model as an explanation for those curves. Students begin to see how accounting for DNA’s elastic resistance to bending and twisting explains the experiments, and are then in a position to evaluate the relative importance of elastic and entropic effects in the original question of DNA packing.
The idea of organizing the syllabus around biological questions can be readily continued for a host of topics. For example, membrane and vesicle mechanics can be motivated by a discussion of encapsulated drug delivery, or the physics of cell adhesion can be motivated by a discussion of the challenges of tissue engineering. The unity of mechanics and statistical mechanics allows those disparate topics to be treated with a limited number of fairly simple ideas.
Go for it
Creating a sustained curriculum change is a big job. But it’s important to meet the challenge: Over the past decade, many physics departments have seen significant erosion in their service teaching and hence their institutional support. We need to explore all possible ways to offer new courses that other departments (and their students) actually want. Moreover, many undergraduate students are basing their choice of graduate school, in part, on the availability of biological physics programs. Soon, undergraduates will choose their majors and high-school students will select their colleges on similar grounds.
But it’s not all about gloomy trends! It’s also exciting to open new doors for students, and gratifying when they respond as they have to our courses. What’s more, we have found teaching biological physics—a course we never took as undergraduates—to be a valuable and fun part of our own professional growth. Some of that excitement has rubbed off on our students, then back onto us, and so on. The prospect of such stimulation is a big part of why we went into academic careers in the first place. So if you find yourself teaching biological physics, remember to enjoy it. And good luck.
We thank Jané Kondev and Rob Phillips for some of the material summarized in this article.
References
1. National Research Council,Bio2010: Transforming Undergraduate Education for Future Research Biologists, National Academies Press, Washington, DC (2003), available at http://www.nap.edu/books/0309085357/html .
2. W. Bialek, D. Botstein, Science 303, 788 (2004) https://doi.org/10.1126/science.1095480
B. Alberts, Cell 92, 291 (1998).3. S. Amador Kane, Am. J. Phys. 70, 581 (2002) https://doi.org/10.1119/1.1473645 .
4. See, for example, B. Ahlborn, Zoological Physics, Springer, New York (2004)
B. Cotterill, Biophysics, Wiley, New York (2002)
K. A. Dill, S. Bromberg, Molecular Driving Forces, Garland, New York (2002)
P. Nelson,Biological Physics: Energy, Information, Life, W. H. Freeman, New York (2004)
B. Nolting, Methods in Modern Biophysics, Springer, New York (2003).5. E. Purcell, Am. J. Phys. 45, 3 (1977);
H. Berg, Random Walks in Biology, 2nd ed., Princeton U. Press, Princeton, NJ (1993) https://doi.org/10.1119/1.10903 .6. See, for example, M. Hoagland, B. Dodson, The Way Life Works, Random House, New York (1995).
7. S. P. Smith et al., Am. J. Phys. 67, 26 (1999) https://doi.org/10.1119/1.19187
J. Bechhoefer, S. Wilson, Am. J. Phys. 70, 393 (2002) https://doi.org/10.1119/1.1445403 .8. R. E. Goldstein et al., http://www.physics.arizona.edu/~gold/ajp.pdf . The Integrative Graduate Education and Research Traineeship program at the University of Arizona is described at http://bmpi.web.arizona.edu .
More about the authors
Ray Goldstein (gold@physics.arizona.edu ) is a professor of physics and applied mathematics and codirector of the quantitative biology initiative of the BIO5 Institute at the University of Arizona in Tucson. Phil Nelson (nelson@physics.upenn.edu ) is a professor of physics at the University of Pennsylvania in Philadelphia and author of the textbook Biological Physics: Energy, Information, Life (W. H. Freeman, 2004). Tom Powers (thomas_powers@brown.edu ) is the James R. Rice Assistant Professor of Solid Mechanics and assistant professor of engineering at Brown University in Providence, Rhode Island.
Raymond E. Goldstein, 1 BIO5 Institute, University of Arizona, Tucson, US .
Philip C. Nelson, 2 University of Pennsylvania, Philadelphia, US .
Thomas R. Powers, 3 Brown University, Providence, Rhode Island, US .





{kind=link}
{kind=link}
{kind=link}
{kind=link}