Mathematical adventures in biology
DOI: 10.1063/1.2709558
The contemplation and resolution of questions at the interfaces of biology, mathematics, and physics promise to lead to a greater understanding of the natural world and to open new avenues for physics. The choice of questions in this article, most of them related to the statistical behavior of biological systems, reflects my own research interests. But it also reflects my belief that some of the unresolved issues in the mathematics of biology are related to the diversity, randomness, variation, and correlations in biology. With luck, physics-based approaches may shed further light on those issues.
The mathematical focus of the research questions I propose complements the public-health focus of the 14 Grand Challenges in Global Health announced in October 2003. 1 Those challenges have realigned health-related research priorities—not only those of the Bill & Melinda Gates Foundation, which initiated the identification of the challenges, but also those of other public and private research funding and public-health bodies. Several of the challenges concern the development of vaccines and drug treatments, topics that invite mathematical analysis.
The following are a few suggestions for mathematical adventures in biology. The first topic concerns the roles of localization, discreteness, and fluctuations in natural biological systems. Cells are not simply well-mixed test tubes, as evidenced by the role of fluctuations in some of life’s processes. The second topic concerns a general question about the mechanism of evolution in biology. Mathematical results might quantify the probability of life as we know it and might be tested in laboratory evolution experiments. The last topic addresses a practical aspect of pathogen evolution. In this case, mathematical results might aid drug or vaccine design, or they might predict the likelihood of future epidemics or the emergence of new pathogens.
The answers to many if not all of the questions suggested in this article lie within the grasp of the physics and biology communities. The answers can be tested experimentally, aid in the interpretation of biological data, and have direct implications for medical practice. They may lead to a more thorough understanding of biology and in all likelihood will lead to an ever-expanding set of new questions.
The roll of fluctuations
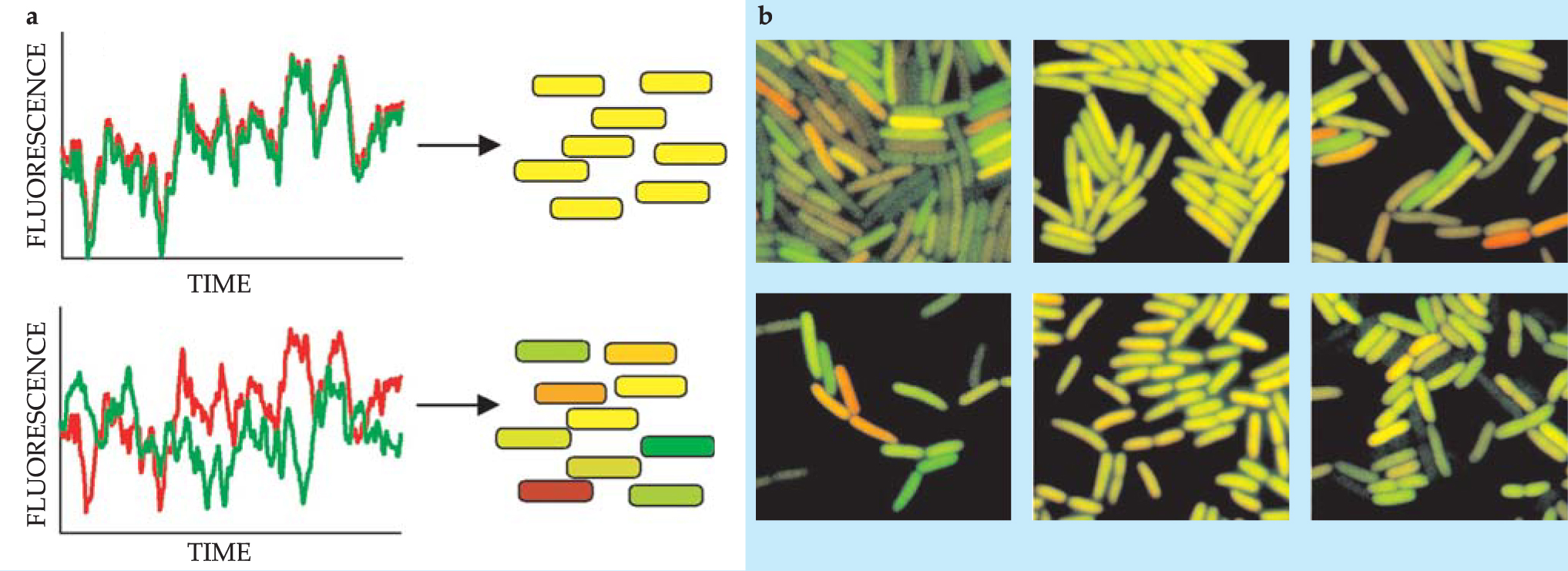
From the cell to the organism, what role does fluctuation play in natural function? Cellular noise arising from the nonuniform distribution of biological molecules in the cell is now appreciated as an interesting physics problem. 2 Figure 1, for example, shows random variations that arise in the concentration of a protein due to stochastic variation of both the environment outside the cell and the biochemical reactions within a cell.

Figure 1. Noise levels are measured with the help of two genes in this experiment with the bacterium Escherichia coli. One gene expresses a green fluorescent protein; another expresses a red fluorescent protein. (a) Fluctuations outside the cell modulate both proteins in the same way (top), so different cells will be the same color. The amount of protein, though, will vary as a function of time. Fluctuations inside the cell modulate the two proteins differently (bottom), so the various cells have different colors in a single experiment. (b) In the six experiments shown, E. coli express differing numbers of the two proteins due to differences in their regulatory mechanisms. Cells expressing fewer proteins have relatively more fluctuations and, therefore, a greater diversity of colors. When does the diversity due to fluctuations in the number of proteins in a cell invalidate the predictions of mean-field theories of cellular function?
(Adapted from M. B. Elowitz et al., ref. 2.)
Many of the molecules of life, such as DNA, mRNA, and enzymes, are present at exceedingly low concentrations—from 1 to 100 molecules per cell. 3 At such low concentrations, the traditional equations of solution chemistry no longer apply, and the stochastic fluctuations associated with the transport and reaction of individual molecules become apparent. At equilibrium, the thermodynamics is that of a small system (see the article by Carlos Bustamante, Jan Liphardt, and Felix Ritort, Physics Today, July 2005, page 43 ). An important detail of biology is that many interesting reactions occur at finite times—that is, when equilibrium has not been established and when dynamics is important. Indeed, one of the differences between much of biology and traditional condensed-matter physics, which has provided useful techniques to the mathematics of biology, is the importance of finite time and finite population size.
A possible way to treat the randomness that arises from reactions at low concentrations is to write a master equation for the process and map that equation onto a bosonic quantum field theory. To date, that approach has been pursued only for the simplest reactions. An alternative, approximate treatment uses mean-field theory, but only in the regions of parameter space where at least one molecule is likely to be present; elsewhere the concentration is simply set to zero. The experiments shown in figure 1 could be described with such a theory, in combination with a description of the statistics of the random extracellular environment.
One might generally ask of biology, under what conditions does the integer constraint on the number of molecules or individuals matter? An integer constraint can induce correlations that traditional mean-field solution-chemistry theory would miss. And traditional theories of population evolution are mean field, so one might expect them to break down at the low concentrations that arise when new drug-resistant strains of bacteria evolve, as pathogens are eradicated, or when species are near extinction.
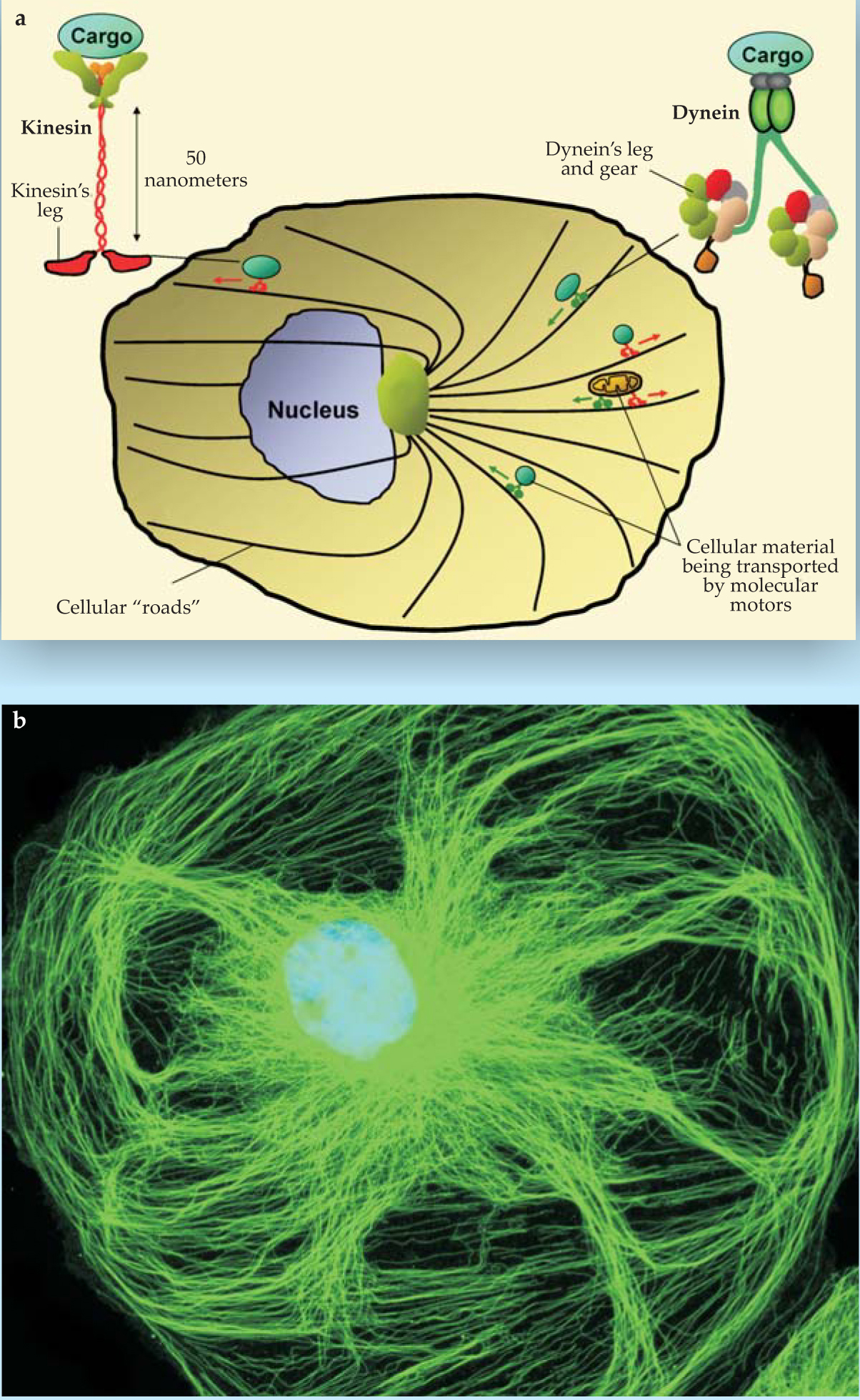
Not only do cellular concentrations of molecules fluctuate thanks to low concentrations, but also the transport of molecules can be active rather than passive. 4 The existence of tracks on which biological molecules are transported within cells (see figure 2) requires a rethinking of traditional solution theory’s assumption that cell transport occurs solely by random diffusive motion. A plethora of questions arise. For example, how do individual microtubules make up the tracks for transport via biological motors, and exactly how do the feet of the motors move during transport? Why does the cell have such diversity of motors, and how may multiple motors act synergistically to transport cargo? What can be said about the organization of the tracks within the cell?

Figure 2. Transport networks facilitate the movement of cellular cargo. (a) A typical cell contains a complex, radially biased network of microtubules, only a small fraction of which is illustrated in this schematic. Also shown here are two of many molecular motors—kinesin, which typically moves cargo toward the cell periphery, and dynein, which typically moves cargo toward the cell’s center. The forces exerted by biological motors are on the order of piconewtons. (b) This image of a rat kangaroo kidney epithelial cell (PtK2) shows the microtubule network, labeled with green fluorescent material. Blue fluorescent dye indicates nuclear DNA. The cell is about 20 μm across.
(
Another complication of the cellular environment is molecular crowding—the relatively high overall concentration of proteins, nucleic acids, and sugars 5 (see the article by Rob Phillips and Steve Quake, Physics Today, May 2006, page 38 ). The cellular volume fraction occupied by such macromolecules is in the range of 5–40%. The tortuous environment they create slows the transport of other large molecules in the cell. Moreover, high levels of molecular crowding can affect cellular reaction rates, by suppressing reactions that create even more crowding and enhancing reactions that alleviate it.
At the level of a human individual one might ask: What role do fluctuation and randomness play in maintaining or relaxing to the natural, healthy physiological state? The healthy state appears to be less a single, stable, fixed point than a rich interplay of complex dynamics. Recent observations suggest that reduction of this natural physiological complexity may be one of the most significant damaging effects of trauma or illness. 6 Perhaps, then, treatment of trauma or illness should introduce a degree of fluctuation or correlated randomness to stimulate the reacquisition of healthy variability. Variation, however, is not typically sought in critical care: Assisted breathing devices deliver identical breaths at a fixed frequency, intravenous or intestinal feedings are given in regular ways, and insulin infusions are typically given at regular intervals.
Can one identify the key features of the nonlinear dynamics of natural physiology that couple to and generate stochastic fluctuations? Is the natural physiological state near a maximum stochastic resonance? In other words, does the level of fluctuation in the system lead to maximum synchronization, responsiveness, or signal detection? Can a fluctuation–dissipation, or response, theory of physiology be developed based on, for example, Peter Hänggi and Harry Thomas’s nonthermal, nonequilibrium, stationary fluctuation–dissipation theory? 7
Surely, some extension of the usual fluctuation-dissipation theorem from equilibrium to nonequilibrium but steady-state systems is necessary to study the physiological state. Because various physiological parameters are measured over time for patients in intensive care units, a fully formed theory might serve as part of the basis for personalized critical care. Many physically interesting responses are to discrete events—for example, feedings, the administration of drugs, or removal from a ventilator—that are routine elements of intensive care. In such cases, the response predicted by the theory can easily be tested. Even though transitions to or from the disease or ill state may be nonlinear, the onset of those transitions may be detectable as violations of the linear theory.
Modular structures
Is it true that evolution of a modular molecular structure allows life to evolve rapidly enough that humans can exist? A modular structure to the molecules of life allows biological information to be stored in pieces. The existence of this modularity means that evolution need not proceed just by changes of one base of the genetic code or movement of one atom or amino acid at a time; rather, functional units can be exchanged among living organisms. For example, as illustrated in figure 3, proteins often comprise almost independent modules, and the genetic information that codes for those modules may be transmitted through evolution. The modular structure of proteins is hierarchical, with identifiable elements at the levels of atoms, amino acids, secondary structures, and domains. Hierarchical elements continue through the levels of proteins, multiprotein complexes, pathways, cells, organs, individuals, and species.

Figure 3. DNA pieces that encode distinct protein modules become organized and concatenated in the course of evolution. The hierarchical organization of biological information appears to allow for rapid evolution in the face of environmental uncertainty. The illustration shows the evolution of the E(Pc)-like protein in yeast (Y), flies (F), and vertebrates (V) to the peregrin protein in flies and humans (H). The insertions of the PHD domains lead to the YPR031w protein in yeast. The insertion of the Br domain leads to the Lin-49 protein in worms (W). The final insertions of the Znf and BMB domains lead to peregrin in flies and humans. Is the hierarchical organization of biological information inevitable?
(Adapted from ref. 16.)

A complementary set of hierarchical mutational events at the genetic level would seem to facilitate the evolution of the molecules of life. And the speed and complexity of evolution do seem to be increasing over time. As indicated by the fossil record, the first single-cell life forms evolved 3.5−4 billion years ago, only 0.5−1 billion years after Earth formed. It took another 2.5−3 billion years for multicellular organisms to appear, but then only 1 billion years for all multicellular species to come into being.
A simplified model of evolution will help to define what I mean by modularity of protein structure and hierarchy of mutational events. Consider a population of individuals undergoing evolution. For example, the individuals may be proteins, and the genetic information that codes for the protein amino-acid sequence may be considered as the evolving variables. Individuals are under selection, which means new ones are produced stochastically. But different individuals replicate at different average rates: Let ri denote the rate at which an individual with sequence i produces another with the same sequence. The population of individuals changes not only by replication but also by stochastic genetic change. In general this random change occurs both during replication and independently of it. For simplicity, consider change independent of replication, and let the mutation matrix μ ij denote the rate for an individual with sequence i to change to one with sequence j. A conventional way to characterize the relatedness of two sequences i and j is to count the number of genetic differences between them. The mutation matrix will surely have nonzero elements connecting closely related sequences with a small number of differences.
Existence of a hierarchy of mutational events implies that the mutation matrix also has large blocks of nonzero elements even when the number of differences is not small. For example, a protein encoded by sequence i may have a whole domain exchanged, which would result in a rather different protein encoded by sequence j. Existence of a modular structure of the protein implies that ri is at least partly decomposable into a linear sum of replication rates associated with pieces of the sequence i, provided that the modules making up the protein can be swapped into or out of a protein without loss of function. Evolution is efficient if distant sequences with large replication rates are connected by nonzero elements of the mutation matrix.
Modularity is not only observed within structures, but also evident in the interactions among the molecules of life. When biologists study those interactions, they are often interested in the topology of the interaction network. In some cases, the network seems rather robust to changes in the values of equilibrium and kinetic constants associated with the interactions between the network’s molecules. 8 Thus, much may be learned about biology by studying the constraints on the interactions. The characteristic structures that arise in the interaction networks also seem modular; figure 4 gives an example. Moreover, investigators have intriguingly suggested that such modularity arises from a pressure to enhance the evolutionary dynamics of the network. 9

Figure 4. The feed-forward loop is one of many network motifs that together form complicated network topologies. Elements X and Y are transcription factors, proteins that activate or repress gene expression. The lower element Z is an operon, a regulated gene that produces a protein. In the feed-forward loop, X influences Y, and the two together influence Z. The online version of this article links to an example of a complicated network that includes feed-forward loops and other motifs: a representation of the entire transcriptional regulation network of Escherichia coli. Can the mathematics describing the constraints on network interactions contribute to biology?
(Adapted from ref. 17.)

Can the advantage of a hierarchical set of mutational events, conjugate to the structural modularity, be shown in a general way? Is it possible to quantify how rapidly evolution may occur by a given set of moves, in the presence of an external environment that includes other species? In special cases, one can show the advantage of a hierarchy, and modular structure is believed by many to be fundamental to evolution. What general statements can be made? Mathematically, one might consider the integral over all possible evolutionary dynamics X, and ask, is the dominant contribution to the system’s response R to environmental change given by those dynamics that are hierarchical, X 0? In symbols,
Assuming that a hierarchy of mutational events does indeed promote rapid evolution, and given that a hierarchy in the natural world is observed, one might ask whether structural modularity is a typical or special case. That is, what is the probability that a modular structure will occur in a general evolutionary system? By asking whether modularity is inevitable and thus inquiring about the probability that life will evolve to evolve via a hierarchy of mutational events, one may understand the structure that is observed in biology without the need to resort to an anthropic or intelligent-design argument. Some string theorists are contemplating a similar question: Given an enormous number of possible universes, is the one we inhabit reasonably likely? An analogy with thermodynamics illustrates the type of answer that researchers are seeking, albeit in a system that seems simpler than biology. Physicists long ago established that in thermodynamics the observed value of energy or density or pressure for a large system is equal to the quantity’s value in the most likely state of the system.
A related question is, what is the underlying pressure for evolution to speed up over time, say by the emergence of modularity and hierarchy? It seems likely that a changing environment selects for adaptable evolutionary frameworks, and competition among different evolutionary frameworks leads to selection for the most efficient dynamics.
Resistance to drugs
How do diseases evolve to escape from vaccines or drugs, and how can scientists design better therapeutics? Pathogens evolve in response to pressure not only from the natural world but also from vaccines or drugs. It has become increasingly apparent, for example, that evolution of drug-resistant bacteria is a problem. 10 Figure 5 quantifies evolving drug resistance for three sample bacteria.

Figure 5. Pathogens evolve in response to other pathogens, the physical environment, and the drugs used to combat pathogens. Emergence of drug-resistant bacteria is rapid and troubling. The graph shows the increasing occurrence of drug resistance in samples collected from intensive-care patients participating in the National Nosocomial Infections Surveillance System, a component of the Centers for Disease Control and Prevention. Three types of bacteria are considered: methicillin-resistant Staphylococcus aureus (blue), vancomycin-resistant Enterococci (green), and fluoroquinolone-resistant Pseudomonas aeruginosa (red). Can biologists learn how to predict and control evolution of drug resistance?
(Source: CDC; see ref. 10.)

Currently, the design of new vaccines and drugs is largely based on the assumption that pathogens evolve in response to therapeutic and immune selection by undergoing minimal changes to the genetic information that encodes them. However, to engineer effective methods of combating viruses, parasites, bacteria, and cancers, scientists may need to anticipate the evolutionary potential of larger changes to the genetic information. For example, the evolution of penicillin resistance occurred by bacterial incorporation of enzymes that degrade the drug and by exchange—sometimes between different bacterial species—of the genetic material that encodes the protein to which penicillin binds. 11 Even influenza and tuberculosis, which were believed to evolve drug resistance largely by an accumulation of single mutations to the genetic sequence, may undergo substantial exchange and recombination between strains. 12
How treatment strategy might depend on disease evolvability, and perhaps how to influence such evolution, is of significant importance to vaccine and drug design efforts. For example, a highly diverse, multicomponent vaccine may be necessary to halt progression and transmission of rapidly mutating viral strains. Conversely, for slowly evolving strains, diversity in the vaccine may simply dilute the conference of protective immunity. What characteristics of a disease need be known to predict evolution? Mutation rate, population size, replication rate, and infectiousness are often considered. 13 Moreover, the network of person-to-person contacts and the variation of immune histories or drug regimens in people also affect pathogen evolution.
Quantitative theories might answer some questions concerning recombination. How useful is recombination to evolution? Are likely recombinations and their utility predictable? The reproduction rate of a pathogen as a function of genetic sequence—the so-called fitness landscape—will be a key component of theoretical descriptions of pathogen evolution. A central issue is the ruggedness of the landscapes on which pathogen evolution occurs. That is, is reproduction rate a gentle function of sequence, or does the fitness landscape have many peaks and valleys? The landscapes seem to be more rugged than originally expected. At one time the consensus of biologists was that organisms evolved drug resistance at a cost, so that if use of the drug were discontinued, reversion to the original, nonresistant strain of the pathogen would occur. Increasingly, reports indicate that the former consensus view is not so; the fitness landscapes of common pathogens include many stable peaks. 14 Can one suggest optimal treatment strategies in light of what is learned about the evolutionary mechanisms and fitness landscapes of pathogens? An ideal goal could be to drive pathogens into regions of low evolvability, where they are most easily eradicated.
At the molecular level, what are the likely escape mutants that lead to new viral strains or new antibiotic-resistant bacteria? To be specific, I’ll consider viruses. The immune system recognizes regions of viral proteins, and it develops the ability to clear the virus from the body based on that recognition. If not completely eradicated from the human (and possibly animal) population, the surviving virus will tend to accumulate mutations in the region of the genetic sequence that codes for those immune-recognizable protein pieces and thus avoid immune recognition. Predicting which mutations are likely in the surviving virus could allow vaccination against potential future strains—the escape mutants—and more effective control of the virus in the population. Similarly, the ability to predict at the molecular level which changes may lead to resistance against a drug can aid in the design of improved drugs or suggest optimal combinations of existing drugs to mitigate the evolution of resistance and eradicate the pathogen.
More dramatic is a point noted by Julie Gerberding, director of the US Centers for Disease Control and Prevention, in a 2005 talk entitled “It’s a Small World for Public Health” she gave at the University of Texas Health Science Center in Houston. Of the last 13 newly observed human viral diseases, 12 are zoonotic—that is, they are animal diseases that have jumped the species barrier to become prevalent in humans. The species-jumping event involves the modification of some poorly understood features of the pathogen, such as transmissibility, viability, and ability to survive human immune attack. At present, it is not possible to predict the propensity for a disease to jump from animals to humans. Learning to make such predictions would be very helpful in planning for future epidemics. One might focus, for example, on the likelihood of events such as the breakout of SARS (severe acute respiratory syndrome) or substantial person-to-person transmission of the H5N1 avian influenza. Do diseases that tend to jump from animals to humans possess generic features? What are the environmental factors that favor the genetic rearrangements leading to such diseases?
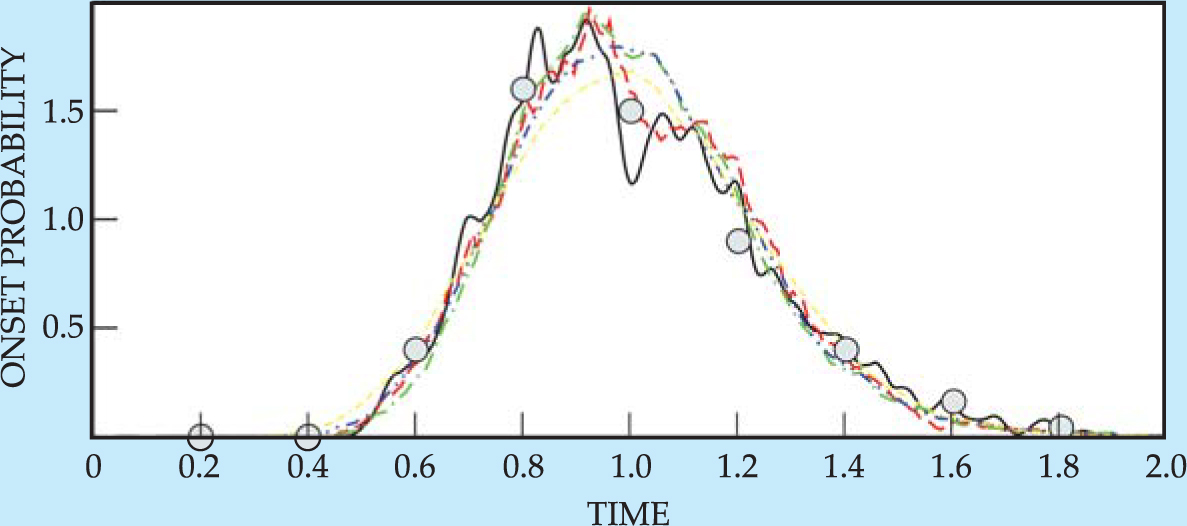
A particularly elegant example in which statistical physics has helped to elucidate disease progression comes from the study of prion neurological diseases by Daniel Cox’s group at the University of California, Davis. The prion protein comes in two forms: the normal PrPC on neuronal surfaces and the aberrant PrPSc. The more hydrophobic PrPSc prions not only form aggregates called plaques but also autocatalyze their formation. Both prion forms are metastable, and which form is globally stable depends on the local molecular environment. By examining the dynamics of the two protein forms diffusing and interconverting in two dimensions, Cox and colleagues found a kinetics of nucleation of the aberrant PrPSc that is in remarkable agreement with onset-time distributions in cattle infected with mad cow disease in the UK. 15 Figure 6 summarizes some of their results.

Figure 6. Incubation time for mad cow disease is amenable to statistical study. The graph shows the onset probability of the disease as a function of time normalized so that the mean incubation time is 1. Filled circles show experimental data; the various curves show simulations for several concentrations of the prion whose aberrant form is associated with the disease.
(Adapted from ref. 15.)
Vaccine and drug design are undergoing a resurgence not only because of their importance to public health and biodefense but also because of the many fascinating associated scientific issues. Statistical physics allows investigation and determination of the fundamental qualitative and quantitative features that govern the interaction between an effective vaccine or drug and the evolving pathogen. Theories of statistical physics may complement the long and difficult process of experimental vaccine and drug development and provide some guidance for those engaged in the endeavor. The successful design of vaccines and drugs must anticipate the evolutionary potential of pathogens in response to therapeutic and immune selection. The addition of disease-specific details to general physical theories of evolution is a promising approach for predicting pathogen plasticity. Infectious agents will continue to evolve unless they are forced down the road to extinction.
I thank Michael Diehl and members of the FunBio team for stimulating discussions. The questions I have discussed in this article are a subset of questions presented first at the 2004 Institute for Complex Adaptive Matter Annual Conference and later at the Frontiers in Physics session at the American Institute of Physics 2005 Industrial Physics Forum.
References
1. H. Varmus et al., Science 302, 398 (2003).
2. M. Thattai, A. van Oudenaarden, Proc. Natl. Acad. Sci. USA 98, 8614 (2001)
M. B. Elowitz et al., Science 297, 1183 (2002)
C. V. Rao, D. M. Wolf, A. P. Arkin, Nature 420, 231 (2002).3. X. S. Xie, J. Yu, W. Y. Yang, Science 312, 228 (2006).
4. R. Mallik, S. P. Gross, Physica A 372, 65 (2006)
R. Mallik, http://www.tifr.res.in/~roop/NaturesNanotech.htm .5. R. J. Ellis, A. P. Minton, Nature 425, 27 (2003).
6. T. G. Buchman, Curr. Opin. Crit. Care 10, 378 (2004). https://doi.org/10.1097/01.ccx.0000139369.65817.b6
7. L. Gammaitoni et al., Rev. Mod. Phys. 70, 223 (1998).
8. G. von Dassow et al., Nature 406, 188 (2000).
9. E. A. Variano, J. H. McCoy, H. Lipson, Phys. Rev. Lett. 92, 188701 (2004).
10. See, for example, the Infectious Diseases Society of America’s“Bad Bugs, No Drugs” webpage, available at http://www.idsociety.org .
11. M. C. J. Maiden, Clin. Infect. Dis. 27, S12 (1998).
12. E. C. Holmes et al., PLoS Biol. 3, e300 (2005)
X. Liu et al., J. Bacteriol. 188, 8169 (2006).13. S. A. Frank, Immunology and Evolution of Infectious Disease, Princeton U. Press, Princeton, NJ (2002).
14. S. Gagneux et al., Science 312, 1944 (2006).
15. A. Slepoy et al., Phys. Rev. Lett. 87, 058101 (2001).
16. E. S. Lander et al., Nature 409, 860 (2001).
17. S. S. Shen-Orr et al., Nat. Genet. 31, 64 (2002).
More about the authors
Michael W. Deem is a professor of physics and astronomy and the John W. Cox Professor in Biochemical and Genetic Engineering at Rice University in Houston, Texas.
Michael W. Deem, Rice University, Houston, Texas, US .





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}